Metilación
La metilación es la adición de un grupo metilo (-CH3) a una molécula.[1] En biología del desarrollo, la metilación es el principal mecanismo epigenético. Aquí la metilación consiste en la transferencia de grupos metilos a algunas de las bases citosinas (C) del ADN situadas previa y continuamente a una guanina (G).[2] Puesto que la metilación es fundamental en la regulación del silenciamiento génico, puede provocar alteraciones en la transcripción genética sin necesidad de que se produzca una alteración en la secuencia del ADN, siendo uno de los mecanismos responsables de la plasticidad fenotípica. También pueden ser metilados los productos de los genes, es decir, las proteínas, regulándose así también su función. En el proceso de metilación intervienen las enzimas conocidas como ADN-metiltransferasas[3].

Hay ocasiones en las que un gen ha de ser desactivado por completo y permanentemente. Para ello se recurre a la metilación del ADN y a la modificación de nucleosomas. Cuando un gen no se expresa, una ADN-metiltransferasa (metilasa) puede metilar citosinas tanto en el promotor, como en la secuencia codificante o en sitios de unión de activadores situados en la posición 5’ respecto del gen. De esta manera, se genera la 5-metilcitosina, la cual por sí sola no es capaz de impedir la unión de la maquinaria de transcripción o de activadores. Para ello se unirán otras proteínas como MeCP2 (proteína que impide la unión de factores de transcripción o bien bloquea el avance de la ARN polimerasa), que reconocen estas metilcitosinas y son además capaces de reclutar al complejo represor Sin3, el cual presenta actividad desacetilasa de histonas.[4][5][6]
Metilación en el desarrollo embrionario
editarEn el desarrollo embrionario temprano, el genoma eucariota es desmetilado. Desde este estado temprano al estado de mórula, se produce la metilación "de novo" de este ADN, modificándose y añadiéndose así la información epigenética de dicho genoma. En el estado de blástula, la metilación se completa. La importancia de la metilación en estos procesos de formación iniciales son fundamentales, ya que se ha comprobado que mutantes en metiltransferasas (que por tanto no pueden metilar su ADN) mueren en estado de mórula.
Recientes estudios han encontrado que en el tejido normal, la metilación de un gen es localizada, principalmente, en la región de codificación, que posee una baja densidad de bases CG. La región , que posee una alta densidad de islas CpG.
Metilación y cáncer
editarEl estado de metilación de algunos genes puede ser usado como un marcador de teratogénesis. En muchas células tumorales la hipermetilación del promotor unida a la pérdida de heterocigosidad en un mismo locus resulta en la pérdida de función de un gen “protector” y comienza a desarrollarse el proceso tumoral. Las reglas que determinan qué genes son metilados durante la patogénesis de un determinado tipo de cáncer son complejas y están aún por descifrar. Sin embargo existen marcadores de metilación del ADN que son muy específicos y que son capaces de detectar muchos tipos de tumores frecuentes. Estos marcadores podrían ser muy útiles en el diagnóstico precoz del cáncer. Por ejemplo, la hipermetilación de la clase pi de la glutatión S-transferasa (GSTP1) parece ser un indicador prometedor de diagnóstico de cáncer de próstata.
Tipos de metilación del ADN
editarExisten dos tipos de metilación del ADN: la de mantenimiento y la de novo:
- La metilación de mantenimiento añade grupos metilo a cadenas de ADN en lugares opuestos a los metilados en la cadena madre, provocando que las moléculas hijas de ADN mantengan un patrón de metilación después de la división celular.[7]
- La metilación de novo del ADN añade grupos metilo en posiciones totalmente nuevas, pudiendo cambiar el patrón de metilación en una región localizada del genoma.[8]
Metilación en el ADN
editarLa metilación de la citosina, normalmente en los CpGs, es una característica común de la regulación epigenética de la expresión génica. La mayoría de los tipos celulares tienen patrones de metilación de dinucleótidos CpG relativamente estables, pero los conocimientos sobre qué CpGs participan en la regulación génica, aún son limitados. Un 70-80% de los CpGs se encuentran metilados. Estos CpGs metilados son detectados mediante la técnica de secuenciación con bisulfito.
Sólo el 21,8% de los CpGs autosómicos tiene una regulación dinámica, la mayoría de ellos distales de puntos de iniciación de la transcripción. Estos CpGs dinámicos, colocalizan con elementos reguladores de genes, particularmente con enhancers y sitios de unión de factores de transcripción (TFBS), lo que permite la identificación de reguladores clave linaje-específicos.
Cabe destacar la importancia de las regiones metiladas diferencialmente (DMRs), las cuales contienen SNPs asociados con enfermedades que son específicas de cada tipo celular.
Aunque en teoría cada CpG es capaz de cambiar su estado de metilación, sólo lo hace una fracción como parte de programas reguladores muy coordinados. Por lo tanto, las DMRs seleccionadas podrían servir como un punto de inicio para guiar nuevas, y más efectivas aproximaciones para capturar la fracción de CpGs más informativa, así como para localizar posibles nuevos elementos reguladores.[9]
Metilación de Purdie
editarLa metilación de Purdie es un método de metilación específica sobre el oxígeno de carbohidratos por el empleo de yodometano y óxido de plata.[10]
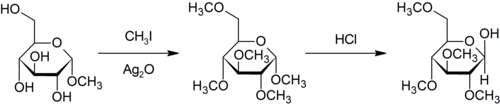
Metilación de Purdie

Véase también
editarReferencias
editar- ↑ «https://www.cancer.gov/espanol/publicaciones/diccionarios/diccionario-cancer/def/metilacion». www.cancer.gov. 2 de febrero de 2011. Consultado el 11 de marzo de 2022.
- ↑ Sánchez, Manuel (2012-05). Farmacología y endocrinología del comportamiento. Editorial UOC. ISBN 9788497884242. Consultado el 9 de diciembre de 2017.
- ↑ «¿Qué es epigenética? ¿Y tú me lo preguntas? Epigenética... eres tú. (Parte II)». Investigación y Ciencia. Consultado el 11 de marzo de 2022.
- ↑ Herráez, Ángel (2012). Texto ilustrado e interactivo de Biología molecular e ingeniería genética. Conceptos, técnicas y aplicaciones en ciencias de la salud. Elsevier. ISBN 978-84-8086-647-7.
- ↑ Watson et al., James D. (2005). Biología Molecular del Gen. Editorial Médica Panamericana. ISBN 84-7903-505-6.
- ↑ Lewin, Benjamin (2008). Genes IX. McGraw Hill. ISBN 970-10-6685-5.
- ↑ Voet, Donald; Voet, Judith G. (2006). Bioquímica. Ed. Médica Panamericana. ISBN 9789500623018. Consultado el 9 de diciembre de 2017.
- ↑ Brown, Terry (30 de junio de 2008). Genomas/ Genome. Ed. Médica Panamericana. ISBN 9789500614481. Consultado el 9 de diciembre de 2017.
- ↑ Ziller MJ. , Gu H, Müller F. , Donaghey J. , Tsai LT. , Kohlbacher O. , De Jager PL. , Rosen ED. , Bennett DA, Bernstein BE. , Gnirke A. , Meissner A. (2013). «Charting a dynamic DNA methylation landscape of the human genome». Nature 500 (7463): 477-81. doi:10.1038/nature12433.
- ↑ Purdie T, Irvine JC (1903). «The alkylation of sugars». J. Chem. Soc. 83: 1021-37. doi:10.1039/CT9038301021.
Enlaces externos
editar Datos: Q518328
Datos: Q518328 Multimedia: Methylation / Q518328
Multimedia: Methylation / Q518328