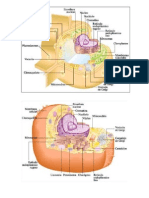
Organoides y Membranas Intracelulares
Organoides y Membranas Intracelulares
Descargar como docx, pdf o txt
También podría gustarte
- Modulo 8Documento18 páginasModulo 8Carlos MegoAún no hay calificaciones
- Componentes EndomembranososDocumento9 páginasComponentes Endomembranososvictor reaAún no hay calificaciones
- Cap. 12 Celula BiologiaDocumento54 páginasCap. 12 Celula BiologiaEmiliano FrancoAún no hay calificaciones
- Modulo 8-04Documento17 páginasModulo 8-04Gustavo RaeleAún no hay calificaciones
- Sistema de EndomembranasDocumento9 páginasSistema de Endomembranasandrea rochaAún no hay calificaciones
- RibosomasDocumento6 páginasRibosomaspakalna80% (5)
- CLASE 6 Reticulo Endoplasmatico MEDICINA 2021 FINALDocumento23 páginasCLASE 6 Reticulo Endoplasmatico MEDICINA 2021 FINALya tu sabeAún no hay calificaciones
- Copia de Sistemas de EndomembranasDocumento8 páginasCopia de Sistemas de EndomembranasMartina AgueroAún no hay calificaciones
- EucariotasDocumento49 páginasEucariotasMohamed AléAún no hay calificaciones
- Sistema de EndomembranasDocumento4 páginasSistema de EndomembranasJosephGabrielAún no hay calificaciones
- Transporte y Relaciones - Membrana PDFDocumento12 páginasTransporte y Relaciones - Membrana PDFBritney Charline AscherAún no hay calificaciones
- EndomembranaDocumento4 páginasEndomembranaJonatham R JiménezAún no hay calificaciones
- MEMBRANAS INTERNAS 1 - TeoríaDocumento25 páginasMEMBRANAS INTERNAS 1 - TeoríaMarino GuerreroAún no hay calificaciones
- Unid 2-Sistemas-de-Endomembranas-docx - GR 6ºDocumento7 páginasUnid 2-Sistemas-de-Endomembranas-docx - GR 6ºMARITZA CORTES PARRAAún no hay calificaciones
- 24 EndomembranaDocumento4 páginas24 EndomembranaFreddy David HerreraAún no hay calificaciones
- Estructura y Función CelularDocumento4 páginasEstructura y Función CelularCarolina Ortiz100% (1)
- Resumen de Biologia Celular - Agosto.2021Documento8 páginasResumen de Biologia Celular - Agosto.2021Michell AndradeAún no hay calificaciones
- 3.membrana Citoplasmica Parte 2 BDocumento11 páginas3.membrana Citoplasmica Parte 2 BMonseAún no hay calificaciones
- Célula AnimalDocumento17 páginasCélula AnimalJUDITH ALONDRA TORRES HERNANDEZAún no hay calificaciones
- TEMA II. Sistemas de Membr. Citoplásmica. Resp. Aeróbica y La Mitoco. Citoesqueleto y Mov. Cel.Documento45 páginasTEMA II. Sistemas de Membr. Citoplásmica. Resp. Aeróbica y La Mitoco. Citoesqueleto y Mov. Cel.BELL TOMLINSONAún no hay calificaciones
- CELULASDocumento4 páginasCELULASNacho PioquintoAún no hay calificaciones
- La CelulaDocumento4 páginasLa CelulaKarolyne RealAún no hay calificaciones
- Unidad Didáctica IvDocumento10 páginasUnidad Didáctica Ivtrasteandopuntocom tpcAún no hay calificaciones
- Citoplasma CelularDocumento19 páginasCitoplasma CelularAlisson Caicedo100% (2)
- Cuestionario Sistema de EndomembranasDocumento4 páginasCuestionario Sistema de EndomembranasLeonora PaelAún no hay calificaciones
- Tarea V Funciones de Cada Una de Las Estructuras CelularesDocumento2 páginasTarea V Funciones de Cada Una de Las Estructuras CelularesEddy Liranzo FañaAún no hay calificaciones
- Informe EditadoDocumento27 páginasInforme EditadoErick SaldañaAún no hay calificaciones
- Tráfico VesicularDocumento55 páginasTráfico VesicularAficionadoAún no hay calificaciones
- Fisio Primer ParcialDocumento34 páginasFisio Primer ParcialFlor SternariAún no hay calificaciones
- Neurociencias ResumenesDocumento15 páginasNeurociencias ResumenesMelany Aldana DAIYOUBAún no hay calificaciones
- 11-Sistema de Endomembranas-Apunte GeneralDocumento19 páginas11-Sistema de Endomembranas-Apunte GeneralTomas Scialfa BenderskyAún no hay calificaciones
- Células Eucariotas y ProcariotasDocumento5 páginasCélulas Eucariotas y ProcariotasFanny CMunvAún no hay calificaciones
- Guia de HistologiaDocumento12 páginasGuia de Histologiayexiimar16Aún no hay calificaciones
- Células EucariotashoDocumento8 páginasCélulas EucariotashoHolger GuamanAún no hay calificaciones
- Celula AnimaDocumento5 páginasCelula Animapo poAún no hay calificaciones
- Organelos CelularesDocumento38 páginasOrganelos Celularesyoselin gonzalezAún no hay calificaciones
- Biología - Química UNED, Tema4Documento6 páginasBiología - Química UNED, Tema4Marta Arias FernandezAún no hay calificaciones
- MitocondriasDocumento6 páginasMitocondriasemmanuelsojuelsilvaAún no hay calificaciones
- Guía de Organelos CelularesDocumento8 páginasGuía de Organelos CelularesCamila Bossio CastillaAún no hay calificaciones
- Modulo III Reticulo Endoplásmicosistema de Endomembranas Con Audio Final 2023Documento29 páginasModulo III Reticulo Endoplásmicosistema de Endomembranas Con Audio Final 2023Madelin BolañosAún no hay calificaciones
- La Membrana PDFDocumento31 páginasLa Membrana PDFCopycentro y Equipos MaryAún no hay calificaciones
- TP Tic Numero 12Documento6 páginasTP Tic Numero 12jl319314Aún no hay calificaciones
- SVC Biología 08Documento7 páginasSVC Biología 08Ptricia OliverosAún no hay calificaciones
- Organelos CitoplasmáticosDocumento24 páginasOrganelos CitoplasmáticosAlbaniBriceñoAún no hay calificaciones
- BiologiaDocumento13 páginasBiologiaHaidith CabreraAún no hay calificaciones
- 03 Premedicina Biologia+Celular+y+Molecular Celula-Organelos+Celulares-transporte+MembranalDocumento9 páginas03 Premedicina Biologia+Celular+y+Molecular Celula-Organelos+Celulares-transporte+Membranalgeorgepiste1Aún no hay calificaciones
- Unidad 11 Organulos Membranosos 2019 2020Documento21 páginasUnidad 11 Organulos Membranosos 2019 2020martaprimenAún no hay calificaciones
- Taller de BiologiaDocumento10 páginasTaller de BiologiaYuveidys Chamorro VergaraAún no hay calificaciones
- Las Entidades Fundamentales de Todo Ser Vivo Son Las CélulasDocumento5 páginasLas Entidades Fundamentales de Todo Ser Vivo Son Las CélulasroximarcontrerasAún no hay calificaciones
- La Célula-Pilar SanchoDocumento12 páginasLa Célula-Pilar SanchoSol MarcosAún no hay calificaciones
- CELULADocumento108 páginasCELULAalejandraAún no hay calificaciones
- Estructura Celula EucarionteDocumento6 páginasEstructura Celula Eucariontebenxtremo99Aún no hay calificaciones
- Biología Unidad 5-Estructura y Función CelularDocumento11 páginasBiología Unidad 5-Estructura y Función CelularJenny MoreiraAún no hay calificaciones
- Capitulo 12 AlbertsDocumento35 páginasCapitulo 12 AlbertsRicardo Cuatlayotl OlarteAún no hay calificaciones
- Apuntes Estructura CelularDocumento12 páginasApuntes Estructura CelularMaria Teresa MolinaAún no hay calificaciones
- Componentes de Endom 184417 Downloadable 5486838Documento11 páginasComponentes de Endom 184417 Downloadable 5486838evelynpapadopulos1Aún no hay calificaciones
- Separata Organelas y Componentes Extracelulares de Una Celula EucariotaDocumento9 páginasSeparata Organelas y Componentes Extracelulares de Una Celula Eucariotalucero liraAún no hay calificaciones
- Los Ribosomas - BiologiaDocumento7 páginasLos Ribosomas - BiologiaMag SosaAún no hay calificaciones
- La CélulaDocumento30 páginasLa CélulaFranco David Castro RamosAún no hay calificaciones
- Taller Informacion en InternetDocumento9 páginasTaller Informacion en InternetDIBL 96Aún no hay calificaciones
- Guia Observación Celular BMS1100Documento10 páginasGuia Observación Celular BMS1100DIBL 96Aún no hay calificaciones
- Guía Huesos Y Sus Funciones: Escuela de SaludDocumento19 páginasGuía Huesos Y Sus Funciones: Escuela de SaludDIBL 96Aún no hay calificaciones
- Ciclo Celular: La Interfase Se Divide en Etapas: G G1, S y G2. Desde El Punto de Vista de La GenéticaDocumento11 páginasCiclo Celular: La Interfase Se Divide en Etapas: G G1, S y G2. Desde El Punto de Vista de La GenéticaDIBL 96Aún no hay calificaciones
- Trabajo QuimicaDocumento4 páginasTrabajo QuimicaDIBL 96Aún no hay calificaciones
- GLANDULAS Y HORMONAS (A)Documento10 páginasGLANDULAS Y HORMONAS (A)DIBL 96Aún no hay calificaciones
- DIGESTIVO AlDocumento10 páginasDIGESTIVO AlDIBL 96Aún no hay calificaciones
- Oportunidades e Ideas 102Documento6 páginasOportunidades e Ideas 102DIBL 96Aún no hay calificaciones
- Trabajo InfoDocumento6 páginasTrabajo InfoDIBL 96Aún no hay calificaciones
- Ecología UnacDocumento83 páginasEcología UnacZAGACETA RODRIGUEZ SHIRLEY ELIZABETHAún no hay calificaciones
- T9 Reino Moneras, Hongos y ProtoctistasDocumento18 páginasT9 Reino Moneras, Hongos y Protoctistasyaniraf007Aún no hay calificaciones
- Las Plantas Alelopaticas PDFDocumento204 páginasLas Plantas Alelopaticas PDFAaron Torres CarhuaninaAún no hay calificaciones
- Arquitectura PeriodontalDocumento53 páginasArquitectura PeriodontalLeo García100% (2)
- Material Complementario de BromatologiaDocumento4 páginasMaterial Complementario de BromatologiaMarlon MAún no hay calificaciones
- Planificacion Cs Naturales 1 6to Nivel CelularDocumento42 páginasPlanificacion Cs Naturales 1 6to Nivel Celularabigail otero100% (1)
- EXAMEN PRÁCTICO II GRUPO C-1 (Parcial 2)Documento4 páginasEXAMEN PRÁCTICO II GRUPO C-1 (Parcial 2)Dilcer Delgado rojasAún no hay calificaciones
- Resumen TraducciónDocumento10 páginasResumen TraducciónCarolina Rivera ReyAún no hay calificaciones
- Diferencia Entre Espermatogénesis y OvogénesisDocumento6 páginasDiferencia Entre Espermatogénesis y OvogénesisAlba DairethAún no hay calificaciones
- Wiley4 TradDocumento28 páginasWiley4 TradJose SalamancaAún no hay calificaciones
- Tesis GlifosatoDocumento7 páginasTesis GlifosatoMaicol PerezAún no hay calificaciones
- Exposicion Bacterio Klebsiella PneumoniaeDocumento14 páginasExposicion Bacterio Klebsiella Pneumoniaediana06jAún no hay calificaciones
- Informe 4 Tincion GramDocumento11 páginasInforme 4 Tincion GramAbril Chavez-Arroyo EnzianAún no hay calificaciones
- Lectura 03 Genética Forense II PDFDocumento15 páginasLectura 03 Genética Forense II PDFDebby DiazAún no hay calificaciones
- Lineamiento Técnico para El Uso de La Ivermectina en Casos Leves de Pacientes Con SARS23 - OKDocumento16 páginasLineamiento Técnico para El Uso de La Ivermectina en Casos Leves de Pacientes Con SARS23 - OKPOPLabMxAún no hay calificaciones
- Vida Útil de Carnes A Temperatura Ambiente y CongelaciónDocumento4 páginasVida Útil de Carnes A Temperatura Ambiente y CongelaciónEverth L TasillaAún no hay calificaciones
- Maria Paula Garcia Garzon Taller Transcripción y Traducción 2Documento9 páginasMaria Paula Garcia Garzon Taller Transcripción y Traducción 2Maria Garcia GarzonAún no hay calificaciones
- Practica de La Cuarta SemanaDocumento7 páginasPractica de La Cuarta SemanaErick Alexis SantiAún no hay calificaciones
- DormiciónDocumento22 páginasDormiciónAntonio OmogrossoAún no hay calificaciones
- Pr-Psa-007 Recuento de Mesofilos Aerobios en AguasDocumento9 páginasPr-Psa-007 Recuento de Mesofilos Aerobios en AguasDianita CortesAún no hay calificaciones
- Leucemia Mieloide CrónicaDocumento5 páginasLeucemia Mieloide CrónicaJessi Torres100% (1)
- Tema 6 PDFDocumento32 páginasTema 6 PDFRODRIGO CACHON VILLAMEDIANAAún no hay calificaciones
- Loque AmericanaDocumento19 páginasLoque AmericanaMauricio NuñezAún no hay calificaciones
- Marco Teorico de Parasitosis TerminadoDocumento23 páginasMarco Teorico de Parasitosis TerminadoVíctor HernándezAún no hay calificaciones
- Unidad 3 Tarea 5Documento15 páginasUnidad 3 Tarea 5Nelson LunaAún no hay calificaciones
- 110-Viruela AviarDocumento6 páginas110-Viruela AviarMAlejandra ContrerasAún no hay calificaciones
- Cientificos y Sus Aportaciones A La GeneticaDocumento3 páginasCientificos y Sus Aportaciones A La GeneticaAngel Adair Salazar Perez100% (3)
- Practica .LipoproteinasDocumento10 páginasPractica .LipoproteinasKarla Liszet Gonzalez HernandezAún no hay calificaciones
- El Aparato Digestivo para Niños de PrimariaDocumento3 páginasEl Aparato Digestivo para Niños de PrimariaanaAún no hay calificaciones
- Clase 1Documento21 páginasClase 1Bri'a M'pmAún no hay calificaciones