Aphididae
Gli Afididi (Aphididae Latreille, 1802) sono una famiglia di circa 2.250 specie di Insetti appartenenti all'Ordine dei Rincoti (sezione Omotteri Sternorrinchi). Sono gli afidi (o pidocchi delle piante) propriamente detti, sebbene questa denominazione si applichi per estensione all'intera Superfamiglia a cui appartengono (Aphidoidea), comprendendo anche gli Adelgidi e le Fillossere.


Descrizione
modificaLa morfologia degli Afididi è rappresentativa della generalità superfamiglia degli Aphidoidea. Pur mantenendo strette analogie con gli Adelgidi e i Fillosseridi, si distinguono da questi per la presenza, nella generalità delle specie, dei caratteristici sifoni addominali e della codicola e per la produzione abbondante di melata. Come tutta la superfamiglia, anche gli afididi sono caratterizzati da un polimorfismo morfologico e funzionale associato al ciclo.
Il corpo è tozzo e piriforme, di dimensioni piccole e di consistenza delicata. La lunghezza varia da 1 a 8 mm, ma in genere non supera i 5 mm. La livrea è poco appariscente, di colore generalmente variabile dal verde al giallo oppure dal bruno al nero, talvolta con tonalità rosate.
Il capo è piccolo e stretto, con antenne relativamente lunghe, composte da 6 articoli nelle forme alate e da 3 nelle attere, tre ocelli e occhi composti provvisti di un processo posteriore recante tre ommatidi. Il rostro è tetramero.
Il torace è poco sviluppato, con ali ben sviluppate nelle forme migranti, del tutto assenti nelle forme meiottere. In relazione alla specificità del ciclo, le forme alate possono essere del tutto assenti all'interno della specie.

C: costa; Pt: pterostigma; Rs: settore radiale; M: rami della media; Cu: rami della cubito.
Le ali sono doppie, trasparenti e membranose. È presente una nervatura longitudinale, che decorre presso il margine costale, ottenuta dalla fusione della subcosta, della radio, della media e della cubito; la nervatura termina con una sclerificazione più o meno allungata (pterostigma), mentre posteriormente si diramano le ramificazioni, per la maggior parte semplici, corrispondenti al settore radiale, ai rami posteriori della media, ai due rami della cubito. Le ali posteriori sono più piccole e presentano una venatura ancora più semplificata. In posizione di riposo vengono tenute verticali sopra l'addome, con il margine costale in basso. Le zampe sono esili e relativamente allungate, di tipo ambulatorio.
L'addome è globoso e marcatamente sviluppato conferendo a questi insetti il tipico aspetto piriforme. Le dimensioni si devono al notevole sviluppo del micetoma e degli organi genitali interni. Sul dorso dell'addome, quasi ai lati, sono in genere presenti, fra gli urotergiti V e VI, due processi, detti sifoni o cornicoli, più o meno lunghi. Per mezzo dei sifoni, gli afidi si difendono dai predatori emettendo una cera fluida che rapprende rapidamente all'aria disturbando il funzionamento dell'apparato boccale[1]. Dai sifoni viene emesso anche un feromone, da parte degli afidi catturati, che mette in allarme gli altri individui della colonia[1]. L'ultimo urite, inoltre, si presenta sottile e allungato, formando una struttura detta codicola, usata per allontanare la melata.
Biologia
modificaRiproduzione
modifica
La riproduzione degli Afididi applica meccanismi che ne esaltano il potenziale riproduttivo tipico degli organismi a strategia r. La riproduzione sessuale è un evento circoscritto ad una fase specifica del ciclo, finalizzata a produrre lo stadio svernante, detto uovo durevole, e garantire perciò la perpetuazione della specie sull'ospite primario e la variabilità genetica. La moltiplicazione è invece affidata alla partenogenesi telitoca, che si svolge nel corso dell'anno producendo un numero indefinito di generazioni di sole femmine e la diffusione delle colonie nell'ambiente.
Alla partenogenesi si associano alcuni fenomeni determinanti sul potenziale biologico di questi insetti: la viviparità, l'inscatolamento delle generazioni e l'endomeiosi.
Gli Afididi, a differenza degli altri Afidi, sono ovipari solo in corrispondenza della riproduzione sessuale, mentre la riproduzione partenogenetica è caratterizzata dalla viviparità. In altri termini, gli individui partenogenetici svolgono il loro sviluppo embrionale all'interno dell'apparato riproduttore della madre e vengono partoriti allo stadio di neanide.

L'inscatolamento delle generazioni (telescoping generations) è un fenomeno che ha importanti ripercussioni sia sulla dannosità degli afidi sia sul loro potenziale riproduttivo. In molte specie le generazioni si annidano all'interno del singolo individuo[2][3].
Dal punto di vista della strategia biologica, questo comportamento fa sì che l'afide investa contemporaneamente le risorse energetiche, acquisite con l'alimentazione, nel duplice obiettivo dell'accrescimento e della riproduzione. Il vantaggio risiede negli intensi ritmi di moltiplicazione e avvicendamento delle generazioni, facendo in modo che le popolazioni degli afidi siano soggette a rapidi incrementi. Nello stesso tempo i fabbisogni nutritivi degli afidi sono notevoli e li rendono perciò organismi dotati di un'elevata voracità. In definitiva questo comportamento intensifica il potenziale di dannosità di questi insetti.
L'endomeiosi è un fenomeno che corregge lo svantaggio, sotto l'aspetto genetico, della riproduzione per partenogenesi: prima della segmentazione dell'uovo partenogenetico si verificherebbero meccanismi di ricombinazione genica, assimilabili al crossing over, fra cromosomi omologhi[4][5]. In definitiva gli Afidi presentano una spiccata variabilità genetica, nonostante la predominanza di meccanismi di riproduzione che sancirebbero la stabilità genetica[6]. Questa effettiva variabilità spiega, almeno in parte, anche la facilità con cui insorgono, nelle generazioni partenogenetiche, fenomeni di resistenza agli insetticidi; naturalmente la partenogenesi e il rapido avvicendamento delle generazioni amplificano la propagazione nella popolazione della resistenza indotta[7].
Polimorfismo
modifica
Il polimorfismo è un altro aspetto notevole della biologia degli Afididi e, più in generale, di tutta la superfamiglia degli Aphidoidea. Questo fenomeno si manifesta principalmente a livello di funzionalità, tuttavia si riscontrano anche polimorfismi morfologici, in particolare in relazione allo sviluppo postembrionale e alla presenza o meno delle ali.
La molteplicità dei cicli e le singole specificità fanno sì che esista una notevole varietà di forme, spesso accompagnata dall'adozione, in letteratura, di terminologie coniate ad hoc. Nella generalità degli Afidi si possono in ogni modo individuare alcune forme biologiche la cui presenza è contestuale a determinate fasi del ciclo; l'esistenza di cicli differenti, più o meno complessi, fa sì che alcune di tali forme scompaiano in una determinata specie, oppure si affianchino ad esse forme intermedie o tipi funzionali particolari.
Facendo riferimento alla riproduzione, le femmine sono distinte nei seguenti tipi funzionali:
- virginopara: nata per partenogenesi, si riproduce a sua volta con lo stesso meccanismo originando una nuova generazione partenogenetica di sole femmine che si riprodurranno per partenogenesi.
- sessupara: nata per partenogenesi, si riproduce a sua volta con lo stesso meccanismo originando però una generazione di individui che si riprodurranno per anfigonia.
- femmina anfigonica: nata per partenogenesi, si riproduce a sua volta per anfigonia, accoppiandosi con un maschio, e deponendo un solo uovo destinato allo svernamento (uovo durevole).
Facendo riferimento al ciclo, si distinguono invece i seguenti tipi morfofunzionali:
- Fondatrice. Femmina nata dall'uovo durevole (anfigonico) e che si riproduce per partenogenesi.
- Fondatrigenie. È una virginopara discendente dalla fondatrice che nasce perciò sull'ospite primario. Può essere attera o alata. Le attere completano il loro ciclo vitale sull'ospite primario generando altre fondatrigenie, le alate si comportano come migranti.
- Migrante. È una fondatrigenie alata che, dopo aver completato lo sviluppo postembrionale, si sposta su un ospite secondario e ivi si riproduce, per partenogenesi, dando inizio ad una serie di generazioni di virginogenie.
- Virginogenie o esule. È una virginopara, discendente da una fondatrigenie, che compie il suo sviluppo sull'ospite secondario. Può essere alata o attera. Le alate migrano su altri ospiti secondari (appartenenti o meno alla stessa specie) propagando linee partenogenetiche parallele di virginogenie.
- Sessupara. Come detto in precedenza, è una femmina che per partenogenesi genera esclusivamente individui destinati alla riproduzione sessuale. La sessupara nasce sempre sull'ospite secondario da una virginopara. L'etologia e la morfologia delle sessupare variano secondo la specie o anche nell'ambito della stessa specie, differenziando alcuni tipi morfofunzionali:
- ginopare: generano esclusivamente femmine; in genere sono alate e si comportano perciò come reimmigranti, riproducendosi sull'ospite primario;
- andropare: generano esclusivamente maschi; in genere sono attere e completano perciò il ciclo vitale sull'ospite secondario;
- anfipare: generano indifferentemente maschi e femmine; sono alate e si comportano perciò come reimmigranti, riproducendosi sull'ospite primario.
- Anfigonico. È un individuo nato per partenogenesi da una sessupara e si riproduce sull'ospite primario accoppiandosi. Si tratta perciò dell'unica generazione in cui si differenziano entrambi i sessi. La morfologia e l'etologia, a prescindere dal sesso, dipende dal comportamento delle sessupare: le femmine sono attere e nascono sempre sull'ospite primario da sessupare reimmigranti; i maschi possono essere atteri oppure alati. In quest'ultimo caso nascono sull'ospite secondario e si comportano perciò come reimmigranti in modo analogo alle sessupare ginopare.
- Iemale. È una virginopara attera caratterizzata da uno sviluppo postembrionale lento in quanto sverna allo stadio di neanide sull'ospite secondario. Dalle iemali si ottengono perciò linee partenogenetiche che alimentano un paraciclo o un anolociclo.
A prescindere dalla relazione con il ciclo biologico, gli individui atteri si possono assimilare a forme neoteniche, in cui permangono caratteri somatici giovanili, di cui i più evidenti sono il meiotterismo e la riduzione della metameria delle antenne. Lo sviluppo postembrionale si svolge in quattro stadi di neanide in cui non si verifica una vera e propria metamorfosi. Gli individui alati, al contrario, subiscono una metamorfosi più spinta in relazione allo sviluppo delle ali e ad una morfologia più complessa. In questi individui, lo sviluppo postembrionale si svolge in quattro fasi: due di neanide, uno di preninfa e l'ultimo di ninfa.
Ciclo biologico
modifica| Per approfondire i concetti e la terminologia di questa sezione vedi Ciclo degli Afidi |
I cicli biologici degli Afididi rientrano nella casistica dei cicli degli afidi sensu lato. In generale hanno una scansione annuale, con alternanza di generazioni polimorfe in cui predominano quelle partenogenetiche.
Il ciclo si completa su un solo ospite (monoico), rappresentato da una specie arborea o arbustiva, oppure si completa su due differenti ospiti (eteroico). In questo caso la biologia dell'afide si complica in quanto il ciclo ha lo scopo di adattarlo a condizioni ambientali particolari: la specie usa differenti ospiti i cui requisiti rispondono a differenti esigenze. Nel ciclo eteroico si distingue un ospite primario, in genere una pianta legnosa, la cui fenologia permette lo svernamento, e un ospite secondario, spesso una pianta erbacea, la cui fenologia si presta a sostenere intensi ritmi di crescita della popolazione. Nel ciclo eteroico la specie sfrutta perciò l'ospite primario al fine di perpetuarsi e quello secondario al fine di moltiplicarsi.
Il ciclo tipo è detto olociclo e si svolge con l'alternanza fra un numero indeterminato di generazioni partenogenetiche e una sola generazione anfigonica. L'olociclo risponde a due differenti esigenze biologiche: la riproduzione sessuale ha lo scopo di perpetuare la specie, permettendone lo svernamento allo stadio di uovo durevole, mentre la partenogenesi ha lo scopo di sostenere la crescita della popolazione. All'olociclo si contrappone l'anolociclo, quando la generazione anfigonica scompare del tutto o diventa rara ed occasionale, oppure si accompagna il paraciclo. In quest'ultimo caso, la specie svolge un regolare olociclo, ma parallelamente sviluppa una linea indipendente di generazioni partenogenetiche che si succedono per più anni sull'ospite secondario, con virginopare iemali che svernano allo stadio di neanide.
Il ciclo, infine, è detto omotopo se si svolge esclusivamente sull'apparato vegetativo aereo (foglie e germogli), eterotopo se alterna generazioni che si sviluppano sulla parte aerea ad altre che invece si insediano sulle radici. Anche in questo caso la differenziazione dei cicli ha lo scopo di adattare la specie a specifiche condizioni: in certi periodi dell'anno, le condizioni sono favorevoli allo sviluppo a spese dell'apparato vegetativo, mentre in altri periodi sono le radici a costituire il substrato trofico per la specie. In genere il ciclo eteroico è anche omotopo, mentre quello eterotopo compare per lo più in specie a ciclo monoico.
Il tipo di ciclo dipende dalla specie ma anche dalle condizioni ambientali. L'olociclo si verifica spesso in regioni caratterizzate da una spiccata stagionalità in quanto la riproduzione sessuale è l'unica che possa permettere lo svernamento e la perpetuazione della specie attraverso l'uovo durevole. La scomparsa delle specie botaniche che possano svolgere la funzione di ospite primario determina la comparsa dell'anolociclo. Questa condizione si verifica spesso con l'importazione di specie provenienti da altre regioni: in genere la regione di nuova introduzione presenta condizioni climatiche più favorevoli e nello stesso tempo una flora che non è in grado di offrire un ospite primario. L'esistenza del paraciclo si verifica invece nelle regioni a clima mite: ad esempio, non poche specie hanno comportamenti differenti secondo la latitudine, svolgendo l'olociclo nelle regioni più a nord e paracicli paralleli all'olociclo nelle regioni più a sud.
In definitiva il determinismo della biologia dell'afide è derivato dal concorso di molteplici fattori, di natura geografica, climatica, ecologica, nutrizionale, genetica e, infine, sociale, con dinamiche estremamente complesse[8]. Ad esempio, il fotoperiodo decrescente la comparsa delle sessupare in tarda estate; nello stesso tempo, nelle fondatrigenie agisce un fattore endogeno che inibisce la comparsa delle sessupare con un effetto decrescente al succedere delle generazioni. Questo comportamento si spiega come adattamento della specie alla stagionalità di un ambiente. La tendenza alla migrazione è invece determinata dal concorso di fattori nutrizionali e sociali: il decadimento della composizione della linfa, con un minore tenore in sostanze azotate, e l'affollamento della colonia si ripercuotono sulla fecondità delle virginopare inducendo una maggiore frequenza della nascita delle alate; in questo modo la specie adotta meccanismi adattativi che le permettono di diffondere le popolazioni su nuovi ospiti, come succede con la migrazione dalle specie arboree a quelle erbacee al passaggio dalla primavera all'estate. Al contrario, nelle colonie in simbiosi con le formiche, queste ultime inibiscono la comparsa delle alate, che tenderebbe a disperdere la colonia con ripercussioni negative sulla disponibilità trofica degli imenotteri.
Nutrizione
modifica
La nutrizione avviene a spese dei succhi cellulari dei tessuti parenchimatici oppure a spese della linfa prelevata dal floema. In quest'ultimo caso l'afide non esercita una suzione, ma è la linfa che risale attraverso l'apparato boccale in ragione della sua pressione. In genere sono attaccati gli organi con i tessuti più teneri, come le foglie, la parte terminale dei germogli, i fiori, ma non mancano casi di specie che si insediano in altre parti della pianta come gli steli lignificati o la base del fusto.
La suzione è sempre preceduta dall'immissione della saliva, che provoca una reazione istologica e fisiologica da parte della pianta. Fra le reazioni istologiche si ha la formazione del canale salivare, ma in molte specie, si verificano alterazioni più evidenti come la formazione di galle o le deformazioni di foglie e germogli. Fra le reazioni fisiologiche si verifica l'aumento dell'attività respiratoria della pianta, la modifica della viscosità della linfa e della sua composizione chimica.
La nutrizione a spese della linfa implica, come nella generalità degli Omotteri, l'adattamento anatomico e fisiologico dell'apparato digerente. In alcuni casi (sottofamiglia Lachninae) gli Afididi dispongono di una rudimentale camera filtrante, ma il funzionamento dell'apparato digerente è fondamentalmente basato sulla simbiosi con i microrganismi del micetoma. I micetomi intervengono anche nel catabolismo dell'azoto, in quanto gli afidi sono privi di tubi malpighiani.
Una diretta della dieta eccessivamente ricca di zuccheri è l'emissione di escrementi liquidi e zuccherini (melata). Le forme libere emettono la melata sotto forma di goccioline che vengono allontanate con bruschi movimenti della codicola. Le forme galligene emettono invece la melata racchiusa in un involucro di cera. La produzione della melata è alla base del rapporto di simbiosi con le formiche.
Etologia
modifica
Gli Afidi sono insetti dotati di scarsa mobilità tendendo alla vita sedentaria e basata fondamentalmente sull'alimentazione. Le alate sono mediocri volatori, ma grazie alle piccole dimensioni e alla leggerezza si lasciano trasportare dal vento anche su grandi distanze.
Di notevole importanza è la tendenza al gregarismo che si manifesta a differenti livelli fino ad una forma rudimentale di eusocialità. Il gregarismo è condizionato da diversi fattori, fra i quali hanno un peso non trascurabile, come si è detto, le interazioni biochimiche tra colonia e ospite, il grado di affollamento nella colonia e la simbiosi con le formiche, oltre a fattori intrinseci legati alla specie.
Dal punto di vista ecologico, il successo della specie risiede nella sua capacità di moltiplicazione; l'individuo destina perciò le proprie risorse energetiche prioritariamente alla riproduzione a scapito dello sviluppo morfogenetico. La massima espressione si ha perciò con la riproduzione partenogenetica associata all'inscatolamento delle generazioni e al mantenimento di una forma sostanzialmente neotenica. La voracità dell'afide, in questo stadio, è tale che le virginopare non cessano di alimentarsi mentre si riproducono. La disponibilità alimentare ha un ruolo essenziale nel condizionare la fecondità delle virginopare e determinare perciò la comparsa delle alate: le trasformazioni istologiche e fisiologiche nelle piante legnose inducono, ad esempio, la comparsa sempre più crescente delle fondatrigenie alate nelle generazioni tardive, spingendo la specie a migrare sulle piante erbacee dalla tarda primavera.

Anche il sovraffollamento produce stimoli ambientali che si riflettono sulla fecondità delle virginopare. In entrambi i casi la ridotta fecondità si manifesta con una maggiore comparsa di individui alati. In questo caso l'obiettivo biologico si sposta dalla moltiplicazione alla diffusione e alla perpetuazione della specie e le risorse energetiche sono preferenzialmente indirizzate allo sviluppo morfogenetico. Le formiche hanno invece un effetto di inibizione nel determinismo delle alate.
Queste complesse relazioni hanno effetti diversi sulle varie specie, per cui si verifica differenze più o meno marcate fra specie che mostrano una spiccata tendenza all'affollamento (es. il Brevicoryne brassicae) rispetto ad altre che hanno una maggiore tendenza alla migrazione (es. il Myzus persicae).
Un aspetto interessante del gregarismo è la dinamica con cui si sviluppa la difesa della specie nei confronti dei suoi nemici naturali. La stessa simbiosi con le formiche produce quale vantaggio principale per gli afidi proprio la protezione attuata dalle formiche contro i predatori: le larve di crisope e coccinellidi vengono letteralmente scacciate dalle operaie, che si oppongono ai loro movimenti nella ricerca delle prede. L'aggregazione, inoltre, stimola meccanismi di difesa rudimentali attraverso segnali chimici: l'afide predato emette dai sifoni il feromone d'allarme, che viene captato dagli altri individui della colonia. La reazione all'emissione del feromone è uno stato di agitazione che pervade la colonia: gli afidi estraggono gli stiletti boccali e si mettono in movimento disperdendosi. Questo comportamento, apparentemente ininfluente per la lentezza dei movimenti, in realtà riduce l'efficienza dei principali predatori in quanto la ricerca della preda si basa sul contatto: infatti il predatore si muove alla cieca e in modo casuale e raggiunge facilmente la preda laddove si ha un maggiore assembramento di afidi[9].
Eusocialità
modifica
L'aspetto più interessante del gregarismo degli Afidi risiede tuttavia nell'eusocialità riscontrata in alcune specie. Le aggregazioni eusociali fra gli insetti, ben note fra gli Imenotteri e gli Isotteri, hanno alcune caratteristiche ricorrenti[10]:
- la stretta associazione fra la comunità e un nido;
- il polimorfismo funzionale finalizzato alla divisione dei compiti;
- la difesa attiva della comunità nei confronti degli aggressori.
L'assenza di uno o più dei suddetti requisiti definisce una forma di aggregazione non sociale: ad esempio le comunità di processionaria del pino non sono eusociali in quanto mancano del secondo e del terzo requisito. A queste forme di aggregazione sono riconducibili quelle della maggior parte degli Aphidoidea. Nelle sottofamiglie Hornaphidinae e Pemphiginae sono state di recente individuate diverse specie con comportamento eusociale, ma gli studi in merito sono ancora in piena evoluzione[10]. Le prime osservazioni, fatte da AOKI e WITHAM, risalgono alla fine degli anni settanta[10][11]. Le ricerche svolte in oltre un ventennio hanno messo in evidenza che in diverse specie, trovate in Giappone, Europa e America, esiste una convergenza evolutiva verso l'eusocialità. I caratteri comportamentali di questi Afididi presentano tutti i requisiti dell'eusocialità: la comunità vive in un nido, rappresentato da una galla o dallo stelo cavo della pianta ospite, e alcuni individui mostrano differenze comportamentali e morfologiche che li destinano a specifici compiti, quali la difesa del nido dall'aggressione da parte dei predatori e la pulizia della galla. A differenza degli altri insetti eusociali, gli afidi hanno tutti corredo cromosomico diploide, sono tutti di sesso femminile ed hanno un patrimonio genetico identico, essendo discendenti di un solo progenitore che si è riprodotto per partenogenesi.

La differenziazione morfofunzionale fa sì che la comunità sia suddivisa in due caste, una specializzata nella riproduzione, l'altra specializzata nella difesa (soldati). Gli afidi soldati, a differenza degli altri individui, sono sterili e allo stadio di neanide, non si alimentano e sono privi di micetomi; la durata della loro vita è dell'ordine di poche settimane, perciò sono destinate a morire precocemente[10][11]. Il compito principale dei soldati è quello di difendere il nido dai potenziali nemici: questi individui stazionano all'esterno del nido e manifestano una spiccata aggressività verso gli insetti predatori e, in alcune specie, anche verso i vertebrati. Gli organi di offesa sono per lo più rappresentati dall'apparato boccale, usato come pungiglione, in grado anche di produrre punture dolorose sui vertebrati, e dalle zampe, usate per lacerare il corpo delle vittime[10]. Oltre alla difesa, è stato riscontrato che i soldati eseguono anche la pulizia della galla, rimuovendo la melata; al momento non si sa, però, se all'interno della casta esiste una suddivisione dei compiti fra difesa e pulizia.
L'eusocialità si manifesta in genere in specie a ciclo eteroico e limitatamente alla fase svolta sull'ospite primario. Il determinismo di casta è ancora in fase di studio, ma si è riscontrata una tendenza a produrre soldati nei periodi dell'anno in cui sono più attivi i predatori. In generale la produzione di soldati aumenta dalla piena estate all'autunno inoltrato e decresce progressivamente nel corso dell'inverno fino alla primavera; contemporaneamente, tende ad aumentare, in primavera il numero di alate[10]. La biologia dell'aggregazione sembra perciò mirata a difendere la colonia finché ci sono i presupposti per la vita sull'ospite primario e quando sono maggiori i rischi per il nido, mentre si orienta alla diffusione della specie, con la comparsa delle migranti, quando ci sono i presupposti per la migrazione sull'ospite primario.
Danni
modifica
Per la loro biologia gli Afididi sono il raggruppamento più dannoso, fatta qualche eccezione, nell'ambito degli Aphidoidea:
- la combinazione fra partenogenesi e viviparità riduce i tempi di sviluppo e intensifica i ritmi di moltiplicazione, rendendo molte specie capaci di produrre pullulazioni in tempi molto brevi;
- il fenomeno dell'inscatolamento delle generazioni rende le virginopare insetti dotati di una notevole voracità in quanto le risorse trofiche sono investite contemporaneamente nello sviluppo e nella riproduzione[2];
- la partenogenesi e il veloce ricambio generazionale facilitano l'insorgenza dei fenomeni di resistenza agli insetticidi;
- molte specie hanno relazioni con virus fitopatogeni di tipo circolativo o di tipo stylet-borne[12], alcuni dei quali di una pericolosità tale da essere più dannosi dello stesso vettore. Basti pensare al virus della Tristeza degli agrumi (Citrus Tristeza Virus), uno stylet-borne trasmesso da alcune specie dei generi Toxoptera e Aphis[13], ritenuta una delle più gravi avversità a carico degli agrumi;
- l'abbondante produzione di melata, oltre a produrre danni collaterali, è la ragione per cui si instaurano facilmente relazioni di simbiosi con le formiche, le quali ne incrementano il potenziale riproduttivo con la loro cura e con la protezione nei confronti dei predatori;
- l'estrema polifagia delle virginopare esuli rende molte specie di facile adattamento a vari contesti produttivi, sfruttando la fase fenologica ottimale di molte piante agrarie, per lo più erbacee.
I danni prodotti dagli Afididi si inseriscono nel quadro generale dei danni causati dagli Omotteri, ma sono spesso amplificati dalla costituzione di colonie numerose e, perciò, sono di gravità tale da inserirli fra gli insetti più dannosi in assoluto per le piante, insieme agli Aleirodidi e alle Cocciniglie:
- alterazioni cromatiche (clorosi) e necrosi fogliari, prodotte per lo più dalla nutrizione parenchimatica;
- deformazioni del lembo fogliare e dell'apice dei germogli, prodotte dalla nutrizione floematica (arricciamenti, arrotolamenti);
- deperimento organico progressivo fino al disseccamento, in genere circoscritto a singole parti della pianta;
- drastica riduzione delle rese;
- trasmissione di virus;
- deprezzamento del prodotto a causa dell'imbrattamento dovuto alla produzione di melata, alle emissioni cerose e ai resti delle mute;
- predisposizione all'insediamento della fumaggine.
Fra i danni specifici prodotti da alcuni gruppi sistematici (es. Pemphiginae) va citate anche la produzione di galle per l'attività auxinica della saliva iniettata.
Difesa
modifica
La difesa dagli afidi è vincolata al grado di biodiversità esistente nell'agrosistema, all'impatto ambientale dell'agrotecnica e al periodo dell'anno in cui si opera. Non meno importante è la biologia della specie: il modo in cui si svolge il ciclo, l'etologia, la mobilità, le dinamiche con cui si determina il rapporto numerico fra migranti e virginopare attere, la polifagia, sono nel complesso fattori che possono condizionare in positivo o in negativo l'impostazione di un'efficace difesa.
La difesa chimica ha una sua efficacia se impostata in modo razionale. In generale si deve tener presente che gli afidi sono insetti poco mobili, che si nutrono esclusivamente aspirando succhi vegetali, che la loro biologia riproduttiva può vanificare gli interventi. Contro le specie che svolgono un olociclo si può intervenire anche con alcuni trattamenti invernali sui fruttiferi, generalmente diretti contro le cocciniglie, ma che hanno secondariamente effetto sulle uova svernanti, tuttavia la lotta preventiva ha scarsa efficacia in quanto, nella generalità dei contesti, le infestazioni sono prodotte soprattutto dall'arrivo delle migranti e dal potenziale riproduttivo delle virginopare. Pochi individui possono infatti generare forti affollamenti localizzati che costituiscono i focolai da cui si propagano successivamente le infestazioni.

La lotta a calendario ha in generale un'efficacia limitata a contesti specifici, come ad esempio il controllo delle prime infestazioni sui fruttiferi nel caso di specie olocicliche eteroiche. Presenta tuttavia diversi effetti collaterali e non è comunque in grado di controllare le infestazioni successive, specie quelle che si svolgono sugli ospiti secondari, sui quali la comparsa degli afidi è del tutto aleatoria. Le strategie più efficaci, perciò, consistono nella lotta guidata o, meglio, nella lotta integrata, che si basano sul principio di intervenire quando la presenza degli afidi supera una soglia di tolleranza. Le soglie di intervento sono generalmente basate sulla frequenza dei germogli infestati, perciò sono facilmente rilevabili contando le colonie presenti in un certo numero di piante.
La scelta dei principi attivi è problematica per alcuni aspetti contrastanti, perciò va ponderata al contesto in cui si opera. Alcuni insetticidi di contatto a largo spettro d'azione hanno in genere un'elevata efficacia, soprattutto per la scarsa mobilità degli afidi. Sono tali i piretroidi, i derivati della nicotina, il rotenone. Il loro impiego si limita però a casi di assoluta necessità per i seguenti motivi:

- a parte alcuni piretroidi, hanno in genere un costo elevato;
- essendo a largo spettro, hanno un forte impatto sull'entomofauna utile;
- hanno uno scarso effetto, se non del tutto nullo, sugli afidi che sfruttano particolari protezioni (galle, deformazioni fogliari, coperture cerose).
In generale l'uso di questi insetticidi va limitato ai casi in cui si verificano infestazioni di colonie che hanno sviluppato una resistenza intrinseca. Nell'ordinarietà si ricorre perciò all'uso di insetticidi endoterapici (translaminari, citotropici e sistemici). Questi principi attivi, infatti, agiscono soprattutto per ingestione, manifestano spesso una selettività intrinseca perché non hanno effetto sugli insetti utili, sono efficaci anche in casi di distribuzione non uniforme e contro colonie protette. In commercio è disponibile una vasta gamma di insetticidi endoterapici specifici o adatti alla lotta contro gli afidi, ma è soggetta ad un forte dinamismo: in genere principi attivi di questo tipo restano in commercio per 10-15 anni, dopo di che vengono ritirati per ragioni normative o perché perdono nel tempo la loro efficacia. L'uso degli endoterapici va limitato per non indurre l'insorgenza della resistenza: un principio attivo va perciò impiegato al massimo 1-2 volte l'anno ed è raccomandata la rotazione fra più principi attivi appartenenti a famiglie chimiche differenti.

Di grande importanza e quasi sempre determinanti sono i fattori naturali di controllo, soprattutto gli insetti entomofagi. Fra i nemici degli afidi, i più importanti sono i seguenti:
- Predatori: larve di Ditteri Sirfidi e di Neurotteri Crisopidi e larve e adulti di Coleotteri Coccinellidi.
- Parassitoidi: larve di Imenotteri Terebranti, la maggior parte inquadrati nella famiglia degli Afelinidi (Chalcidoidea) e nella sottofamiglia degli Aphidiinae (Braconidae).
Nel controllo naturale, gli entomofagi sono in genere efficaci in tarda primavera, fino a determinare la scomparsa delle infestazioni in estate negli agrosistemi ben mantenuti. Con la lotta integrata può essere perciò necessario ricorrere ad uno o due trattamenti mirati contro le infestazioni precoci, lasciando ai fattori naturali il proseguimento del controllo. In alternativa si deve invece applicare la lotta biologica con metodo inondativo, ricorrendo a lanci di crisope. In caso di infestazioni localizzate e fortemente disperse, i migliori risultati si ottengono tuttavia con i sirfidi, in grado di disperdersi meglio nell'agrosistema.
Sistematica
modificaSulla sistematica degli Aphididae vi sono due diverse interpretazioni. La prima, ricorrente soprattutto in vecchi schemi tassonomici, suddivide la superfamiglia degli Aphidoidea in due sezioni, distinguendo gli afidi ovipari (Adelgidae e Phylloxeridae) da quelli vivipari (esclusa la riproduzione sessuale). Quest'ultima, denominata Aphidoidea vivi-ovipara, corrisponde agli Aphididae sensu lato, ma viene suddivisa in famiglie, fra le quali si annoverano anche gli Aphididae sensu stricto.
Una classificazione più moderna raggruppa invece gli afidi vivi-ovipari nell'unica famiglia degli Aphididae, suddivisa in sottofamiglie. L'inquadramento sistematico degli Aphididae sensu lato è anch'esso controverso: un'interpretazione considera gli Aphididae l'unica famiglia compresa negli Aphidoidea, separando gli Adelgidi e i Fillosseridi in una superfamiglia a parte (Adelgoidea o Phylloxeroidea); l'altra, adottata in questa sede, raggruppa le tre famiglie nella stessa superfamiglia, quella degli Aphidoidea. In ogni modo la suddivisione interna è ancora incerta in quanto in continua evoluzione e non del tutto condivisa. La tendenza è quella di scorporare alcuni gruppi (secondo gli Autori considerati al rango di famiglia o sottofamiglia) in più sottofamiglie, ciascuna rappresentata da pochi generi. Diverse fonti considerano a tutt'oggi il raggruppamento dei Drepanosiphinae molto più vasto, comprendendo in esso undici delle sottofamiglie sottoelencate. Ne consegue che gli inquadramenti sistematici adottati anche nella letteratura più recente sono piuttosto controversi. Fra le sottofamiglie citate nelle pubblicazioni compaiono le seguenti:
- Anoeciinae
- Aphidinae
- Calaphidinae
- Chaitophorinae
- Drepanosiphinae
- Greenideinae
- Hormaphidinae
- Israelaphidinae
- Lachninae
- Lizeriinae
- Macropodaphidinae
- Mindarinae
- Myzocallidinae
- Meophyllaphidinae
- Pemphiginae (syn. Eriosomatinae)
- Phloeomyzinae
- Phyllaphidinae
- Pterastheniinae
- Pterocommatinae
- Saltusaphidinae
- Spicaphidinae
- Taiwanaphidinae
- Tamaliinae
- Thelaxinae
Di queste, la più importante per numero di specie è quella degli Aphidinae, comprendente oltre 2000 specie. Le altre comprendono un numero variabile da poche decine a poche centinaia di specie.
Note
modifica- ^ a b Tremblay, p. 105.
- ^ a b Tremblay, pp. 106-108.
- ^ P. Kindlmann, A.F.G. Dixon, Developmental Constraints in the Evolution of Reproductive Strategies: Telescoping of Generations in Parthenogenetic Aphids, in Functional Ecology, vol. 3, n. 5, 1989, pp. 531-537. URL consultato il 9 ottobre 2008. Abstract.
- ^ G. Cognetti, Endomeiosis in parthenogenetic lines of Aphids, in Cellular and Molecular Life Sciences, vol. 17, n. 4, 1961, pp. 168-169, DOI:10.1007/BF02160362, ISSN 1420-682X. URL consultato il 1º dicembre 2020 (archiviato dall'url originale il 29 aprile 2016). Abstract.
- ^ K.A. Shufran, D.C. Peters, J.A. Webster, Generation of clonal diversity by sexual reproduction in the greenbug, Schizaphis graminum, in Insect Molecular Biology, vol. 6, n. 3, 2003, pp. 203-209, DOI:10.1046/j.1365-2583.1997.00174.x.
- ^ Tremblay, pp. 105-106.
- ^ Tremblay, p. 117.
- ^ Tremblay, pp. 111-112.
- ^ Giorgio Celli, Stefano Maini, Giorgio Nicoli. La fabbrica degli insetti. Padova, Franco Muzzio Editore, 1991. ISBN 88-7021-569-5.
- ^ a b c d e f g Judd.
- ^ a b Tremblay, p. 167.
- ^ Vedi il paragrafo Relazioni tra afidi e virus in Aphidoidea.
- ^ (EN) 00.017.0.01.008. Citrus tristeza virus, su ICTVdB Descriptions, International Committee on Taxonomy of Viruses. URL consultato il 15 ottobre 2008 (archiviato dall'url originale il 9 settembre 2006).
Bibliografia
modifica- Sebastiano Barbagallo (coordinamento). Gli afidi e le colture agrarie. REDA, 1985.
- (EN) Roger L. Blackman, Victor F. Eastop. Aphids on the World's Herbaceous Plants and Shrubs. 1ª. Chichester, John Wiley & Sons, 2006. ISBN 0-471-48973-5.
- (EN) Roger L. Blackman, Victor F. Eastop. Aphids on the World's Crops: An Identification and Information Guide. 2ª. Chichester, John Wiley & Sons, 2000. ISBN 0-471-85191-4.
- Michael Chinerey. Guida degli insetti d'Europa: atlante illustrato a colori. Padova, Franco Muzzio editore, 1998. ISBN 88-7413-025-2.
- A. E. Douglas, Nutritional interactions in insect-microbial symbioses: Aphids and their symbiotic Bacteria Buchnera, in Annual review of entomology, vol. 43, 1998, pp. 17-37, DOI:10.1146/annurev.ento.43.1.17, ISSN 0066-4170. URL consultato il 17 ottobre 2008. Abstract.
- Sergio Gengotti, Il controllo degli Afidi nell'agricoltura "Bio" (PDF), in Agricoltura, vol. 36, n. 6, pp. 99-100. URL consultato il 17 ottobre 2008 (archiviato dall'url originale il 12 giugno 2009).
- (EN) Tim Judd, The Behavioral Ecology of the Social Aphids, su colostate.edu, Colorado State University. URL consultato il 13 ottobre 2008 (archiviato dall'url originale il 5 settembre 2008).
- George C. McGavin. Insetti, ragni e altri artropodi terrestri. Guida fotografia a oltre 300 specie di tutto il mondo. F. Garibaldi (trad. it.). 3ª ed. Fabbri, 2008. ISBN 88-451-7574-X.
- (PT) Ângelo Moreira da Costa Lima. XXIII. Homópteros in Insetos do Brasil. Tomo 2. Escola Nacional de Agronomia, 1940.
- Aldo Pollini. Manuale di entomologia applicata. Bologna, Edagricole, 2002. ISBN 88-506-3954-6.
- Gabriele Pozzi. Insetti. Giunti Editore, 1998. ISBN 88-09-03532-1.
- Antonio Servadei, Sergio Zangheri, Luigi Masutti. Entomologia generale ed applicata. Padova, CEDAM, 1972.
- Ermenegildo Tremblay, Entomologia applicata, Volume II Parte I, 1ª ed., Napoli, Liguori Editore, 1981, ISBN 978-88-207-1025-5.
Altri progetti
modifica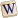 Wikizionario contiene il lemma di dizionario «Afididi»
Wikizionario contiene il lemma di dizionario «Afididi» Wikimedia Commons contiene immagini o altri file su Afididi
Wikimedia Commons contiene immagini o altri file su Afididi Wikispecies contiene informazioni su Afididi
Wikispecies contiene informazioni su Afididi
Collegamenti esterni
modifica- (EN) aphid, su Enciclopedia Britannica, Encyclopædia Britannica, Inc.
- (EN) Aphididae, su Fossilworks.org.
- (EN) The Aphidoidea Web Page, su sel.barc.usda.gov, Systematic Entomology Laboratory, US Department of Agriculture. URL consultato il 17 ottobre 2008 (archiviato dall'url originale il 2 ottobre 2008).
- (EN) David L. Stern, Nancy A. Moran, Aphidoidea, su tolweb.org, The Tree of Life Web Project. URL consultato il 17 ottobre 2008 (archiviato dall'url originale il 13 dicembre 2013).
- (EN) Family Aphididae, in Australian Faunal Directory, Australian Government, Department of the Environment, Water, Heritage and the Arts. URL consultato il 28 febbraio 2009.
- (EN) APHIDIDAE (a family of aphids), su BioImages: The Virtual Field-Guide (UK). URL consultato il 17 ottobre 2008 (archiviato dall'url originale l'8 luglio 2008).
- Fabio Stoch, Family Aphididae, in Checklist of the Italian fauna online version 2.0, 2003. URL consultato il 17-10-2008.
- (EN) Aphididae, su Forestry Images: Forest Health, Natural Resources & Silviculture Images, The Bugwood Network & USDA Forest Service. URL consultato il 17 ottobre 2008.
- (FR, EN) Aphididae, Callaphididae, Pemphigidae, su HYPP Zoologie en ligne, Institut national de la recherche agronomique. URL consultato il 10 maggio 2007.
- (EN) Superfamily: Aphidoidea, su BugGuide. URL consultato il 17 ottobre 2008.
- (EN) Taxon details: Aphidoidea, in Fauna Europaea version 2.6.2, Fauna Europaea Web Service, 2013. URL consultato il 17-10-2008.
- (EN) ITIS Standard Report Page: Aphidoidea, in Integrated Taxonomic Information System. URL consultato il 17-10-2008.