
Diatomácea: diferenças entre revisões
| Linha 68: | Linha 68: | ||
A maior parte das diatomáceas apresenta uma [[frústula]] composta por duas [[valva]]s (ou [[Teca (biologia)|tecas]]), que se sobrepõem e se encaixam como as partes de uma [[placa de Petri]]. A teca maior (ou superior) é designada por ''[[epiteca]]'', a menor (ou inferior) é designada por ''hipoteca''. A região de encontro entre as duas valvas é designada por ''pleura'' ou ''fissura pleural''. |
A maior parte das diatomáceas apresenta uma [[frústula]] composta por duas [[valva]]s (ou [[Teca (biologia)|tecas]]), que se sobrepõem e se encaixam como as partes de uma [[placa de Petri]]. A teca maior (ou superior) é designada por ''[[epiteca]]'', a menor (ou inferior) é designada por ''hipoteca''. A região de encontro entre as duas valvas é designada por ''pleura'' ou ''fissura pleural''. |
||
A [[frústula]] pode possuir ornamentações diversas, entre as quais os poros, que estão arranjados em padrões diferentes entre as espécies, não contendo [[sílica]] nessas regiões. A frústula aparece delicadamente ornamentada com relevos que formam desenhos variados e perfeitamente simétricos, dando lugar a dois tipos de diatomáceas, as que apresentam simetria radial e a que apresentam de simetria lateral. De acordo com a simetria corporal, podem ser definidos dois grupos: as diatomáceas penadas (simetria bilateral ou |
A [[frústula]] pode possuir ornamentações diversas, entre as quais os poros, que estão arranjados em padrões diferentes entre as espécies, não contendo [[sílica]] nessas regiões. A frústula aparece delicadamente ornamentada com relevos que formam desenhos variados e perfeitamente simétricos, dando lugar a dois tipos de diatomáceas, as que apresentam simetria radial e a que apresentam de simetria lateral. De acordo com a simetria corporal, podem ser definidos dois grupos: as diatomáceas penadas (simetria bilateral ou [[Pennales]]) e as diatomáceas cêntricas (simetria radial ou [[Centrales]]). |
||
As frústulas das diatomáceas, por terem densidade superior, sedimentam em meio aquático (marinho e outros) por gravidade quando morre e é digerida a célula, dando origem, quando em grandes quantidades, a [[rocha sedimentar|rochas sedimentares]] como as [[diatomite]]s e [[moronite]]s. |
As frústulas das diatomáceas, por terem densidade superior, sedimentam em meio aquático (marinho e outros) por gravidade quando morre e é digerida a célula, dando origem, quando em grandes quantidades, a [[rocha sedimentar|rochas sedimentares]] como as [[diatomite]]s e [[moronite]]s. |
||
=== Excreção de mucilagem=== |
=== Excreção de mucilagem=== |
||
{{AP|Mucilagem}} |
{{AP|Mucilagem}} |
||
Revisão das 14h17min de 16 de março de 2020
| Bacillariophyta Diatomáceas | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
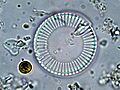
 Diatomáceas marinhas (imagem de microscopia óptica). | |||||||||||||||
| Classificação científica | |||||||||||||||
| |||||||||||||||
| Subgrupos[1] e classes[2] | |||||||||||||||
| |||||||||||||||
| Sinónimos | |||||||||||||||
| |||||||||||||||







Diatomáceas (também frequentemente designadas por Diatomea, Diatomeae, Bacillariophyta ou Bacillariophyceae sensu lato) é a designação dada em biologia e taxonomia a um numeroso e biodiverso agrupamento taxonómico de algas unicelulares que na sua presente circunscrição taxonómica contém mais de 20 000 espécies extantes. O grupo caracteriza-se pela presença de um tipo específico de parede celular, designado por frústula, composta por sílica opalina. Constitui um dos tipos mais comuns de fitoplâncton, assumindo na maioria dos ambientes marinhos e lacustres função dominante entre os produtores (organismos autotróficos) que formam a base da cadeia alimentar.[3][4][5] A maioria das espécies ocorre na camada eufótica dos mares de águas frias, mas são muito numerosas as espécies de água doce e comuns as espécies que crescem à superfície do solo. Apesar da diversidade dos habitats onde ocorrem diatomáceas, a vasta maioria das espécies flutua nos mares e lagos, representando uma parcela importante do fitoplâncton e sendo responsável pela libertação de cerca de 70% do oxigénio atmosférico da Terra. Contudo, algumas espécies produzem uma mucilagem adesiva e vivem presas à superfície de organismos marinhos, como macroalgas, moluscos, crustáceos, tartarugas e baleias e em habitats como a superfície de troncos de árvores, superfície de rochas expostas e mesmo sobre as folhas de plantas.[4]
Descrição
As diatomáceas são organismos unicelulares, ainda que em muitas das espécies coexistam indivíduos isolados com formas coloniais do tipo filamentoso ou em cadeias celulares (p. ex. Fragillaria), em forma de leque (p. ex. Meridion), estruturas em ziguezague (p. ex. Tabellaria) ou formas estreladas (p. ex. Asterionella). Apesar de unicelulares, as diatomáceas são organismos que chegam a atingir 2 milímetros de comprimento.
Uma característica específica e distintiva deste tipo de algas é apresentarem as suas células rodeadas por uma parede celular única, constituída por dióxido de silício (SiO2), ou sílica opalina (dióxido de silício hidratado), designada por frústula. As frústulas são verdadeiras carapaças minúsculas, exibindo uma ampla variedade na sua forma, mas geralmente compostas por duas partes assimétricas separadas por uma linha divisória bem marcada, característica que dá o nome ao grupo.
Na maioria das espécies, a frústula é formada por duas partes encaixadas (algo como uma caixa redonda com tampa), com saliências, depressões e poros que permitem contacto da membrana plasmática com o meio exterior. Essa estrutura confere a muitas espécies de diatomáceas um aspecto iridescente e brilhante. A iridescência das carapaças siliciosas, associada aos pigmentos fotossintéticos presentes, dá às algas varias colorações, indo do dourado ao castanho-esverdeado. As principais substâncias de reserva das diatomáceas são óleos (acumulados em oleoplastos, que em muitas espécies contribuem para facilitar a flutuação.
Em certas regiões do fundos marinho ou lacustres as carapaças de diatomáceas acumularam-se ao longo de centenas de milhar ou mesmo milhões de anos, formando camadas rochosas compactas conhecidas como diatomitos (ou terras de diatomáceas quando sejam rochas pouco consolidadas). As terras de diatomáceas são utilizadas desde a Antiguidade Clássica europeia como material de construção, geralmente misturadas com cal. Alguns exemplos de grandes obras construídas com terras de diatomáceas, e que ainda subsistem, são os aquedutos de Roma, os portos de Alexandria e o Canal de Suez. Por serem constituídos por carapaças vitrificadas muito pequenas, os diatomitos apresentam granulosidade finíssima, sendo por isso frequentemente usados como matéria-prima de polidores e também para confecção de filtros e isolantes.
O registo fóssil conhecido sugere que as diatomáceas se originaram durante ou depois do período Jurássico Inferior, apesar dos primeiros restos corpóreos conhecidos estarem datados do Paleogeno. As comunidades de diatomáceas são uma ferramenta usada frequentemente para a vigilância das condições ambientais, nomeadamente da qualidade da água e do seu estado trófico, e no estudo das alterações climáticas.
Morfologia
O grupo apresenta enorme biodiversidade, com centenas de milhar de espécies e formas extantes, para além daquelas que apenas são conhecidas do registo fóssil. As suas paredes celulares de sílica, as frústulas, apresentam grande diversidade de formas, muitas delas extraordinariamente intrincadas e ornamentadas, que tornam as diatomáceas um dos grupos de seres vivos microscópicos que mais atenção tem despertado. Esta biodiversidade traduz-se em mais de 250 géneros, não contando os extintos, conhecendo-se mais de 20 00 espécies de diatomáceas, com alguns autores a estimarem que o número total seja superior a 100 000 espécies.[6]
As diatomáceas pertencem a um grande grupo de protistas designado por Stramenopiles (ou Heterokontophyta no anterior sistema de classificação), que inclui conjuntamente espécies autotróficas (como as Phaeophyceae, as algas-castanhas) e heterotróficas (como os fungos aquáticos do grupo Oomycetes, os oomicetes). As diatomáceas são na sua vasta maioria autotróficas, mas algumas espécies são, por adaptação secundária, heterotróficas ou mixotróficas.
Os cloroplastos de coloração amarelo-acastanhado das diatomáceas são típicos dos heteroncontes, apresentando clorofilas a, c1 e c2 e pigmentos acessórios como o β-caroteno, a fucoxantina e outras xantofilas, entre as quais a neofucoxantina, a diatoxantina e a diadinoxantina.[7] Os cloroplastos apresentam a estrutura típica dos plastos dos heterocontófitos, sendo organelos com quatro membranas plasmáticas, o que aponta para uma origem por endocitobiose secundária.[8][9]
As reservas de alimento são armazenadas como carboidratos ou lípidos, que para além de servirem de reserva, contribuem para flutuabilidade. Os materiais de reserva preferenciais são a crisolaminarina e gotas de lípidos, em oleoplastos que para além da sua função metabólica auxiliam no controlo da flutuação ao permitirem variar a densidade da célula.[10]
Os indivíduos adultos deste grupo não apresentam flagelos, embora estes estejam presentes nos gâmetas e geralmente com a típica estrutura heteroconte, diferenciando-se contudo por carecerem dos mastigonemas características dos restantes grupos de Heterokonta. Em geral, as diatomáceas não apresentam capacidade de movimento próprio, embora algumas espécies possam controlar o seu deslocamento pela contracção repetida da rafe, única região flexível dada a rigidez da parede celular siliciosa que constitui a frústula.
As diatomáceas tendem a formar cadeias de células, as quais ajudam a manter a flutuação das células, essencial para que possam permanecer na zona eufótica, coincidente com as águas superficiais, dada a dependência destas organismos da fotossíntese. Algumas espécies regulam activamente sua flutuabilidade com lípidos intracelulares, reduzindo a sua densidade e assim evitando a subsidência.
Várias espécies são endossimbiontes de extrema importância em cnidários e em muitas das espécies que formam os recifes de coral, formando parte importante das zooxantelas. Estes endossimbiontes fotossintéticos, que em muitos casos são responsáveis pela coloração dos organismos hospedeiros, fornecem capacidade fixadora de energia solar e são indispensáveis à sobrevivência das espécies heterotróficas a que estão associados.
Algumas espécies apresentam bioluminescência, enquanto outras produzem toxinas que podem penetrar na cadeia trófica através de peixes e de organismos filtradores.
Podem reproduzir-se sexuada ou assexuadamente. Os espécimes adultos carecem, usualmente, de flagelos, ainda que estes estejam presentes nos gâmetas, apresentando a estrutura típica dos heterocontófitos, diferenciando-se contudo pela ausência de mastigonemas, as fibrilhas que estão presentes na maioria dos grupos de heterocontes.
Composição da parede celular
Apesar de ser geralmente microscópicas, algumas espécies de diatomáceas podem alcançar os 2 mm de comprimento. As células das diatomáceas estão contidas dentro de uma única parede celular de silicato (frústula) composta por duas tecas (ou valvas) separadas. A parede celular das diatomáceas, que forma a frústula, é composta, em peso, por cerca de 95% de sílica biogénica e 5% de compostos orgânicos.
A sílica biogénica da parede celular é sintetizada intracelularmente por polimerização de monómeros de ácido silícico. Este material é secretado para o exterior da célula produzindo, em conjugação com os cerca de 5% em peso de material proteico e glúcidos, a estrutura da parede celular.
A maior parte das diatomáceas apresenta uma frústula composta por duas valvas (ou tecas), que se sobrepõem e se encaixam como as partes de uma placa de Petri. A teca maior (ou superior) é designada por epiteca, a menor (ou inferior) é designada por hipoteca. A região de encontro entre as duas valvas é designada por pleura ou fissura pleural.
A frústula pode possuir ornamentações diversas, entre as quais os poros, que estão arranjados em padrões diferentes entre as espécies, não contendo sílica nessas regiões. A frústula aparece delicadamente ornamentada com relevos que formam desenhos variados e perfeitamente simétricos, dando lugar a dois tipos de diatomáceas, as que apresentam simetria radial e a que apresentam de simetria lateral. De acordo com a simetria corporal, podem ser definidos dois grupos: as diatomáceas penadas (simetria bilateral ou Pennales) e as diatomáceas cêntricas (simetria radial ou Centrales).
As frústulas das diatomáceas, por terem densidade superior, sedimentam em meio aquático (marinho e outros) por gravidade quando morre e é digerida a célula, dando origem, quando em grandes quantidades, a rochas sedimentares como as diatomites e moronites.
Excreção de mucilagem
Devido a ausência de flagelos, as diatomáceas apresentam uma motilidade que é produzida pela excreção de mucilagem, produzida no citoplasma e libertada através das ornamentações corporais.[11][7]
Entre as Pennales, devido à existência de uma rafe (fenda longitudinal, de onde também é libertada mucilagem), a movimentação ocorre por adesão do substrato, causada pela mucilagem. Entre as Centrales, o deslizamento é proporcionado por canais de sílica, designados por pórtulas, que secretam mucilagem através de uma abertura que se estende do citoplasma até a face externa da teca.[11][7]
Multiplicação e reprodução
As diatomáceas reproduzem-se essencialmente por multiplicação celular vegetativa: as duas tecas da frústula separam-se, a célula divide-se, e cada uma das tecas que a constituíam assume a função de epiteca (teca ou valva maior), reconstituindo uma hipoteca (teca ou valva menor).
Como consequência deste processo de multiplicação, ocorre uma diminuição progressiva do tamanho das células-filhas que recebem a hipoteca, para fazerem dessa a sua epiteca. Quando um tamanho mínimo é atingido, a reprodução sexuada surge para restabelecer o tamanho normal da espécie, por meio de um zigoto que é designado por auxósporo[7]. Em alguns casos, a reprodução sexuada é desencadeada por mudanças nas condições físicas do ambiente ou na disponibilidade de nutrientes.[11]
As células vegetativas são diploides pelo que a produção de gâmetas ocorre por meiose.
Distribuição e ecologia
A enorme biodiversidade do grupo e o cosmopolitismo da sua distribuição, aliada à enorme biomassa que agregadamente representa e à sua capacidade fotossintética, fazem das diatomáceas um dos agrupamentos biológicos mais importantes a nível planetário, constituindo parte substancial da base das cadeias tróficas das regiões oceânicas.
Distribuição
As diatomáceas constituem um agrupamento biológico com distribuição cosmopolita, com presença comum nos mais diversos habitats aquáticos ou com elevada disponibilidade de humidade de todas as regiões do planeta. Embora a vasta maioria das espécies ocorra na camada eufótica dos mares de águas frias, o grupo está presente em todos os oceanos, nas massas de água doce, na superfície do solo e numa grande variedade de superfícies húmidas (como a face de rochedos, os troncos de árvores e mesmo a superfície das folhas das plantas) em locais com humidade atmosférica elevada.[11]
Apesar dessa diversidade de habitats, a maioria das espécies são organismos marinhas pelágicos de vida livre presentes na camada eufótica das águas oceânicas. Contudo as diatomáceas são também de ocorrência frequente entre as espécies fitobênticas, ocupando a superfície da interface entre o substrato sedimentar e a água nos fundos marinhos e lacustres pouco profundos e bem iluminados (já que estes organismos necessitam da luz solar para viver).[11] Algumas espécies produzem uma mucilagem adesiva e vivem presas à superfície de organismos marinhos, como macroalgas, moluscos, crustáceos, tartarugas e baleias.
As diatomáceas são especialmente importantes nos oceanos, onde se estima que contribuam com mais de 25% da produção primária planetária,[11] ao assegurarem 45% da produção primária oceânica.[12] O conjunto das diatomáceas é responsável pela libertação de cerca de 70% do oxigénio atmosférico da Terra.
As diatomáceas habitam em todos os tipos de corpos de agua, desde as regiões polares até às latitudes mais baixas, embora as regiões polares e subpolares contenham relativamente poucas espécies em contraste com a diversidade do biota das regiões temperadas e tropicais.[13] Contudo, apesar das regiões tropicais exibirem a maior quantidade de espécies, as maiores populações de diatomáceas ocorrem nas águas marinhas das regiões polar e temperada, onde a disponibilidade de nutrientes é maior.[14]
Algumas espécies formam parte do conjunto de endossimbiontes fotossitéticos, conhecidos por zooxantelas, presentes em organismos marinhos, assumindo extrema importância nos recifes de coral.
Em conclusão, apesar da grande diversidade de habitats que ocupam, a maioria das espécies ocorre na camada eufótica dos mares de águas frias ricas em nutrientes, onde representam uma parcela importante do fitoplâncton. Apesar disso, é grande a diversidade de espécies, mesmo em habitats confinados, como a superfície de troncos de árvores, as faces de rochas expostas e mesmo sobre as folhas de plantas.[4]
Ecologia
As diatomáceas são microorganismos fotossintetizadores que formam parte do fitoplâncton. Como colonizadores, as diatomáceas distinguem-se por estarem presentes em qualquer corpo de água, seja marinho, dulceaquícola ou mesmo sobre superfícies húmidas em ambientes terrestres.
Algumas espécies podem ser consideradas entre os extremófilos, ocorrendo em ambientes onde existem condições extremas de temperatura ou de salinidade. Também ocorrem em formas que depende da interacção com outros organismos, como é o caso do epifitismo das diatomáceas sobre as cianofíceas.
A distribuição espacial do fitoplâncton marinho está limitada, tanto horizontal como verticalmente, essencialmente pela disponibilidade de radiação solar e de nutrientes. Em consequência, as populações de diatomáceas, sendo estas produtores primários, estão limitadas às águas eufóticas onde existam nutrientes disponíveis, sendo nestas frequentemente factor limitante a disponibilidade de micronutrientes como o ferro.
História geológica
As diatomáceas surgiram provavelmente no início do Triássico, como resposta à libertação de diversos nichos ecológicos após a Extinção Permo-Triássica. Os primeiros fósseis de diatomáceas, que correspondem às primeiras formas silicificadas, surgem no Jurássico e diversificaram-se bastante a partir de então. No Cretácico a flora de diatomáceas é muito abundante e há exemplos de formações rochosas - os diatomitos - constituídas quase inteiramente de carapaças de diatomáceas.
Também conhecidas por algas douradas, porque possuem uma carapaça rígida de tom dourado, as diatomáceas são muito comuns em oceanos e mares. Algumas regiões do Nordeste brasileiro possuem depósitos naturais de suas carapaças, chamados de diatomitos, material utilizado, por exemplo, na fabricação de produtos para polimento, cosméticos e tijolos.
Importância econômica
As frústulas de sílica das diatomáceas vêm se acumulando ao longo de milhões de anos nos sedimentos oceânicos, formando uma substância fina e porosa que é conhecida como diatomito. Essa substância é usada como abrasivo para polir prata, como material isolante e filtro.
Nos campos de petróleo de Santa Maria da Califórnia existe um depósito subterrâneo de diatomito com cerca de 900 m de espessura e, próximo a Lompoc, Califórnia, mais de 270.000 t de diatomito são retiradas anualmente para o uso industrial[11].
Classificação biológica
Baseando-se em características estruturais e ultraestruturais, tanto quanto na fisiologia desses organismos (i.e., mecanismos de reprodução assexuada e sexuada), Round et al. (1990) desenvolveram um sistema de classificação (o qual se encontra descrito logo abaixo). Entretanto, desde então, alguns gêneros foram criados e/ou ressuscitados. Alguns desses gêneros encontram-se posicionados na classificação abaixo, o que pode diferir do sistema, Ipsis literis, de Round e colaboradores.
Divisão BACILLARIOPHYTA
Classe COSCINODISCOPHYCEAE
Subclasse THALASSIOSIROPHYCIDAE
Ordem THALASSIOSIRALES Glezer & Makarova 1986.
Família Thalassiosiraceae Lebour 1930
Thalassiosira, Planktoniella, Porosira, Minidiscus, Bacteriosira.
Família Skeletonemataceae Lebour 1930,
Sketletonema, Detonula
Família Stephanodiscaceae Glezer & Makarova 1986
Dyscostella, Cyclotella, Cyclostephanos, Stephanodiscus, Mesodictyon, Pleurocyclos, Stephanocostis.
Família Lauderiaceae ( Schütt) Lemmermann 1899,
Lauderia
Subclasse COSCINODISCOPHYCIDAE
Ordem CHRYSANTHEMODISCALES Round, 1990.
Família Chrysanthemodiscaceae Round 1978
Chrysanthemodiscus
Ordem MELOSIRALES Crawford, 1990.
Família Melosiraceae Kützing 1844,
Melosira, Druridgea
Família Stephanopyxidaceae Nikolaev
Stephanopyxis
Família Endictyaceae Crawford, 1990.
Endictya
Família Hyalodiscaceae Crawford, 1990.
Hyalodiscus, Podosira
Ordem PARALIALES Crawford, 1990.
Família Paraliaceae Crawford 1988
Paralia, Ellerbeckia
Ordem AULACOSEIRALES Crawford, 1990.
Família Aulacoseiraceae Crawford, 1990.
Aulacoseira, Strangulonena
Ordem ORTHOSEIRALES Crawford, 1990.
Família Orthoseiraceae Crawford, 1990.
Orthoseira
Ordem COSCINODISCALES Round & Crawford, 1990.
Família Coscinodiscaceae Kützing 1844
Coscinodiscus, Palmeria, Stellarima, Brughtwellia, Craspedodiscus
Família Rocellaceae Round & Crawford, 1990.
Rocella
Família Aulacodiscaceae (Schütt) Lemmermann 1903
Aulacodiscus
Família Gossleriellaceae Round, 1990.
Gossleriella
Família Hemidiscaceae Hendey 1937 emend Simonsen 1975
Hemidiscus, Actinocyclus, Azpeitia, Roperia
Família Heliopeltaceae H. L. Smith 1872
Actinoptychus, Glorioptyschus, Lepidodiscus
Ordem ETHMODISCALES Round, 1990.
Família Ethmodiscaceae Round 1990.
Ethmodiscus
Ordem STICTOCYCLALES Round, 1990.
Família Stictocyclaceae Round 1978
Stictocyclus
Ordem ASTEROLAMPRALES Round & Crawford, 1990.
Família Asterolampraceae H L. Smith 1872
Asterolampra, Asteromphalus
Ordem ARACHNOIDISCALES Round, 1990.
Família Arachnoidiscaceae Round, 1990.
Arachnoidiscus
Ordem STICTODISCALES Round & Crawford, 1990.
Família Stictodiscaceae (Schütt) Lemmermann 1899
Stictodiscus
Subclasse BIDDULPHIOPHYCIDAE
Ordem TRICERATIALES Round & Crawford, 1990.
Família Triceratiaceae (Schütt) Lemmermann 1899
Triceratium, Odontella, Lampriscus, Sheshukovia, Pseudoauliscus, Eupodiscus, Pleurosira, Amphitetras, Cerataulus, Auliscus
Família Plagiogrammaceae De Toni 1890
Pladiogramma, Glyphodesmis, Dimeregramma, Dimerogrammopsis
Ordem BIDDULPHIALES Krieger 1954.
Família Biddulphiaceae Kützing 1844
Biddulphia, Biddulphiopsis, Hydrosera, Isthmia, Trigonium, Terpsinoë, Pseudotriceratium
Ordem HEMIAULALES Round & Crawford, 1990.
Família Hemiaulaceae Heiberg 1863
Hemiaulus, Eucampia, Climacodium, Cerataulina, Trinacria, Abas, Briggera, Pseudorutilaria, Keratophora, Kittonia, Strelnikovia, Riedelia, Baxteriopsis, Sphyntolethus, Ailuretta
Família Bellerocheaceae Crawford, 1990.
Bellerochea, Subsilicea
Família Streptothecaceae Crawford, 1990.
Streptotheca, Neostreptheca
Ordem ANUALES Round & Crawford, 1990.
Família Anaulaceae (Schütt) Lemmermann 1899
Anaulus, Eunotogramma, Porpeia
Subclasse LITHODESMIOPHYCIDAE
Ordem LITHODESMIALES Round & Crawford, 1990.
Família Lithodesmiaceae Round, 1990.
Lithodesmium, Lethodesmioides, Ditylum
Subclasse CORETHROPHYCIDAE
Ordem CORETHRALES Round & Crawford, 1990.
Família Corethraceae Lebour 1930
Corethron
Subclasse CYMATOSIROPHYCIDAE
Ordem CYMATOSIRALES Round & Crawford, 1990.
Família Cymatosiraceae Hasle, von Stosch & Syvertsen 1983
Cymatosira, Campylosira, Plagiogrammopsis, Brockmanniela, Minutocellus, Leyanella, Arcocellulus,Papiliocellulus, Extubocellulus
Família Rutilariaceae De Toni 1894
Rutilaria, Syndetocystis
Subclasse RHIZOSOLENIOPHYCIDAE
Ordem RHIZOSOLENIALES Silva 1962.
Família Rhizosoleniaceae De Toni 1890
Rhizosolenia, Proboscia, Pseudosolenia, Urosolenia, Guinardia, Dactiliosolen
Família Pyxillaceae (Schütt) Simonsen 1972
Pyxilla, Gladius, Gyrodiscus, Mastogonia, Pyrgupyxis
Subclasse CHAETOCEROTOPHYCIDAE
Ordem CHAETOCEROTALES Round & Crawford, 1990.
Família Chaetocerotaceae Ralfs in Pritchard 1861
Chaetoceros, Gonioceros, Bacteriastrum
Família Acanthocerataceae Crawford, 1990.
Acanthoceros
Família Atteyaceae Round & Crawford, 1990.
Attheya
Ordem LEPTOCYLINDRALES Round & Crawford, 1990.
Família Lepocylindraceae Lebour 1930
Leptocylindrus
Classe FRAGILARIOPHYCEAE
Subclasse FRAGILARIOPHYCIDAE
Ordem FRAGILARIALES Silva 1962 .
Família Fragilariaceae Greville 1833
Fragilaria, Centronella, Asterionella, Staurosirella, Staurosirella, Staurosira, Pseudostaurosira, Punctastriata, Fragilariaforma, Martyana, Diatoma, Synedra, Ctenophora, Neosynedra, Tabularia, Catacombas, Hyalosynedra, Opephora, Trachysphenia, Thalassioneis, Falcula, Pteroncola, Asterionellopsis, Bleakeleya, Podocystis, Meridion
Ordem TABELLARIALES Round, 1990.
Família Tabellariaceae Kützing 1844
Tabellaria, Tetracyclus, Oxyoneis
Ordem LICMOPHORALES Round, 1990.
Família Licmophoraceae Kützing 1844
Licmorpha, Licmosphenia
Ordem RHAPHONEIDALES Round, 1990.
Família Rhaphoneidaceae Forti 1912
Rhaphoneis, Diplomenora, Delphineis, Neodelphineis, Perissonoë, Sceptroneis
Família Psammodiscaceae Round & Mann, 1990.
Psammodiscus
Ordem ARDISSONIALES Round, 1990.
Família Ardissoneaceae Round, 1990.
Ardissonea
Ordem TOXARIALES Round, 1990.
Família Toxariaceae Round, 1990.
Toxarium
Ordem THALASSIONEMATALES Round, 1990.
Família Thalassionemataceae Round, 1990.
Thalassionema, Thalassiothrix, Trichotoxon
Ordem RABDONEMATALES Round & Crawford, 1990.
Família Rhabdonemataceae Round, 1990.
Rhabdonema
Ordem STRIATELLALES Round, 1990.
Família Striatellaceae Kützing 1844
Striatella, Microtabella, Grammatophora
Ordem CYCLOPHORALES Round & Crawford, 1990.
Família Cyclophoraceae Round & Crawford, 1990.
Cyclophora
Família Entopylaceae Grunow 1862
Entopyla, Genphyria
Ordem CLIMACOSPHENIALES Round, 1990.
Família Climacospheniaceae Round, 1990.
Climacosphenia, Synedrosphenia
Ordem PROTORAPHIDALES Round, 1990.
Família Protoraphidaceae Simonsen 1970
Protoraphis, Pseudohimantidium
Classe BACILLARIOPHYCEAE
Subclasse EUNOTIOPHYCIDAE
Ordem EUNOTIALES.
Família Eunotiaceae Kützing 1844
Euniotia, Actinella, Semiorbis, Desmogonium
Família Periniaceae (Karsten) Topachevs’kyj & Oksiuk 1960
Peronia
Subclasse BACILLARIOPHYCIDAE
Ordem LYRELLALES D.G.Mann, 1990
Família Lyrellaceae D. G. Mann, 1990.
Lyrella, Petroneis
Ordem MASTOGLOIALES D.G.Mann, 1990
Família Mastogloiaceae Mereschkowsky 1903
Aneumastus, Mastogloia
Ordem DICTIONEIDALES D.G.Mann, 1990
Família Dictyoneidaceae D. G. Mann, 1990.
Dictyoneis
Ordem CYMBELLALES D.G.Mann, 1990
Família Rhoicospheniaceae Chen & Zhu 1983
Rhoicosphenia, Campilopyxis, Cuneolus, Gomphoseptatum, Gomphonemopsis
Família Anomoeoneidaceae D. G. Mann, 1990.
Anomoeoneis, Staurophora
Família Cymbellaceae Greville 1833
Placoneis, Cymbella, Brebissonia, Encyonema, Gomphocymbella, Encyonopsis, Pseudoencyonema, Navicymbulla, Afrocymbella, Gomphocymbellopsis, Delicata, Cymbellopsis, Cymbopleura
Família Gomphonemataceae Kützing 1844
Gomphonema, Didymosphenia, Gomphoneis, Reimeria, Gomphopleura
Ordem ACHNANTHALES Silva 1962.
Família Achnanthaceae Kützing 1844
Achnanthes
Família Cocconeidaceae Kützing 1844
Cocconeis, Campyloneis, Anorthoneis, Bennetella, Epipellis
Família Achnanthidiaceae D. G. Mann, 1990.
Achnanthidium, Eucocconeis, Planothidium
Ordem NAVICULALES Bessey 1907
Sub-ordem Neiidineae D. G. Mann, 1990.
Família Berkeleyaceae D. G. Mann. 1990.
Parlibellus, Berkeleya, Climaconeis, Stenoneis
Família Cavinulaceae D. G. Mann. 1990.
Cavinula
Família Cosmioneidaceae D. G. Mann. 1990.
Cosmioneis
Família Scholioneidaceae D. G. Mann. 1990.
Scolioneis
Família Diadesmidaceae D. G. Mann, 1990.
Diadesmis, Luticola
Família Amphipleuraceae Grunow 1862
Frickea, Amphipleura, Frustulia, Cistula
Família Brachysiraceae D. G. Mann, 1990.
Brachysira
Família Neidiaceae Mereschkowsky 1903
Neidium
Família Scoliotropidaceae Mereschkowsky 1903
Scoliopleura, Scoliotropis, Biremis, Progonoia, Diadema
Sub-ordem Sellphorineae D. G. Mann, 1990.
Família Sellaphoraceae Mereschkowsky 1902
Sellaphora, Fallacia, Rossia, Caponea
Família Pinnulariaceae D. G. Mann, 1990.
Pinnularia, Diatomella, Oestrupia, Dimidiata
Sub-ordem Phaeodactilineae J. Lewin 1958
Família Phaeodactylaceae Silva 1962
Phaeodactylum
Sub-ordem Diploneidineae D. G. Mann, 1990.
Família Diploneidaceae D. G. Mann, 1990.
Diploneis, Raphidodiscus
Sub-ordem Naviculineae Hendey 1937
Família Naviculaceae Kützing 1844
Navicula, Trachyneis, Pseudogomphonema, Seminavis, Rhoikoneis, Haslea, Cymatoneis
Família Pleurosigmataceae Mereschkowsky 1903
Pleurosigma, Toxonidea, Donkinia, Gyrosigma, Rhoicosigma
Família Plagiotropidaceae D. G. Mann, 1990.
Plagiotropis, Stauropsis, Pachyneis
Família Stauroneidaceae D. G. Mann, 1990.
Stauroneis, Craticula
Família Proschkiniaceae D. G. Mann, 1990.
Proschkinia
Ordem THALASSIOPHYSALES D.G.Mann, 1990
Família Catenulaceae Mereschkowsky 1902
Catenula, Amphora, Undatella
Família Thalassiophysaceae D. G. Mann, 1990.
Thalassiophysa
Ordem BACCILARIALES Hendey 1937.
Família Bacillariaceae Ehremberg 1831
Bacillaria, Hantzschia, Psammodictyon, Tryblionella, Cymbellonitzschia, Gomphonitzschia, Gomphotheca, Nitzschia, Denticula, Denticulopsis, Fragilariopsis, , Cylindrotheca, Simonsenia, Cymatonitzschia, Perrya
Ordem RHOPALODIALES D.G.Mann, 1990
Família Rhopalodiaceae (Karsten) Topachevs’kyj & Oksiyuk 1960
Epithemia, Rhopalodia, Protokeelia
Ordem SURIRELLALES D.G.Mann, 1990
Família Entomoneidaceae Reimer in Patrick & Reimer 1975
Entomoneis
Família Auriculaceae Hendey 1964
Auricula
Família Surirellaceae Kützing 1844
Hydrosilicon, Petrodictyon, Plagiodiscus, Stenopterobia, Surirella, Campylodiscus, Cymatopleura
Referências
- ↑ Christiaan Hoek et al. 1995, Algae: An Introduction to Phycology. Cambridge University Press
- ↑ Wolfgang Frey (Ed.) 2015, Syllabus of Plant Families - A. Engler's Syllabus der Pflanzenfamilien Part 2/1: Photoautotrophic eukaryotic Algae. ISBN 978-3-443-01083-6
- ↑ Chapman, A. D. (2009). Numbers of living species in Australia and the world Arquivado em 28 de setembro de 2015, no Wayback Machine..
- ↑ a b c Round, F. E. and Crawford, R. M. (1990). The Diatoms. Biology and Morphology of the Genera, Cambridge University Press, UK.
- ↑ Canter-Lund, H. and Lund, J.W.G. (1995). Freshwater Algae, Biopress Limited. ISBN 0 948737 25 5.
- ↑ F. E. Round, R. M. Crawford, D. G. Mann, Diatoms: Biology and Morphology of the Genera. Cambridge University Press, 1990.
- ↑ a b c d REVIERS, BRUNO (2006). Biologia e filogenia das algas. Porto Alegre: Artmed. 160 páginas
- ↑ Peter Kroth & Heinrich Strotmann, "Diatom plastids: Secondary endocytobiosis, plastid genome and protein import". Physiologia Plantarum, Vol. 107 (1999), issue 1, pp. 136-141.
- ↑ Andreas K. Michels, Norbert Wedel & Peter G. Kroth, "Diatom Plastids Possess a Phosphoribulokinase with an Altered Regulation and No Oxidative Pentose Phosphate Pathway". Plant Physiology: 137 (3), Mar 2005.
- ↑ Yoshiaki Maeda, Daisuke Nojima, Tomoko Yoshino & Tsuyoshi Tanaka, "Structure and properties of oil bodies in diatoms", Phil. Trans. R. Soc. B, 372 (2016).
- ↑ a b c d e f g RAVEN, PETER (2007). Biologia Vegetal. RJ: Guanabara Koogan
- ↑ D. G. Mann (1999), "The species concept in diatoms". in "Phycological reviews" 18. Phycologia 38:437–495.
- ↑ Salinas-Márquez, F.M., Flores-Trujillo, J.G., Helenes, J., Téllez-Duarte, M.A., Aranda-Manteca, F.J., (2016). «Paleoecología y cronoestratigrafía de las diatomeas del Miembro Los Indios en la Mesa La Misión, del Mioceno de Baja California, México» (PDF). Boletín de la Sociedad Geológica Mexicana. 68 (3): 537-552
- ↑ Mann, D. G. (1999). The species concept in diatoms. Phycologia 38, 437-495.
Galeria
-

-
-

-
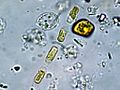
 Melosira sp.
Melosira sp. -
-
-








-

-
-
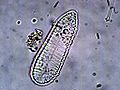
-
 Cymbella sp.
Cymbella sp. -
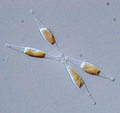
 Licmophora sp.
Licmophora sp.




