zyxwv
zyxwvutsrqponmlk
zyxwvutsrqponmlkjihg
zyxwvutsrqponm
zyxwvutsrqponmlk
zyxwvutsrqponm
zyxwvutsrqpo
00??-1042 78 0901-0751102000
Jourtrol u/Mulrrorhr,,ll,rn. Vol. 31. pp 751 753
Pcrgsmon Press Ltd 1978 Printed in Great Britain
0 International Society lor Neurochcmistry Ltd.
SHORT COMMUNICATION
Subcellular localization of growth hormone
release inhibiting hormone in rat
hypothalamus, cerebral cortex, striatum and thalamus
(Receiivd 9 January 1978. Accepted 23 March 1978)
GROWTHhormone release inhibiting hormone (GH-RIH)
is present in high concentration in the hypothalamus, but
has a wide extrahypothalamic distribution in the CNS
(BROWSTEINet al., 1975). Although immunocytologic
studies have revealed GH-RIH immunoreactivity in nerve
endings in the external layer of the median eminence as
et al., 1975). viswell as in hypothalamic axons (HBKFELT
ualisation of GH-RIH immunoreactivity in extrahypothalamic CNS has been unsuccessful using these techniques.
We have used subcellular fractionation of CNS homogenates as a more sensitive means of studying localisation
in these areas employing a sensitive and specific radioimmunoassay for GH-RIH (KRONHEIM
et a\.. 1976). We describe in this paper our findings on the subcellular distribution of GH-RIH immunoreactivity in the hypothalamus,
striatum. thalamus and cerebral cortex of the rat, and show
the concentration of this immunoreactivity to reside largely
in the synaptosome fraction.
30 min whilst controls of identical composition were incubated at 4°C to assess non-specific choline or noradrenaline uptake. Uptake was terminated by rapid cooling in
an ice bath following which unincorporated tracer was
largely removed by centrifuging the mixture at 10,000g
for 5 min at 4°C. The resultant pellet. resuspended by
gentle homogenisation in 0.32 M-sucrose. was then applied
to a linear continuous density gradient of 0.32 M-1.5 M-SUCrose prepared in 30 ml cellulose centrifuge tubes (Beckman
Instruments) and centrifuged at 75,000 g for 90 min in an
zyxwvutsrqp
MATERIALS AND METHODS
Male Long-Evans rats (20G300g) housed under constant conditions were rapidly decapitated between 09.00
and 10.00 h, the brains removed to an ice tray and dissected, as previously described (KRONHEIM
er al., 1976),into
the striatum, thalamus, hypothalamus and cerebral cortex.
In each experiment, the areas from 5 to 10 rats were pooled
and a 107, w/v homogenate prepared in 0.32~-sucrose
using a Perspex pestle in a glass homogeniser (clearance
0.25 mm) (ALDRIGEe f al., 1960) at 900 rev./min. Homogenisation was performed as described by B ~ N N E T &
T
EUWARUSON
(1975).
G H - R I H profile and neurotransrnirter uptake on continuous sucrose density gradient. In initial experiments. the
profile of homogenate immunoreactive GH-RIH was compared to that of '"C choline and 'H noradrenaline uptake
by tissue fractionated on a continuous sucrose density gradient. The homogenate was allowed to warm slowly to
room temperature. One millilitre was then added to 5 ml
of Krebs bicarbonate buffer (pregassed with 95:: O,, 5:/,
C 0 2 ) containing. in addition, glucose (10 mM). adenosine
triphosphate (0.45 mM), sodium pyruvate (10 mM), ascorbic
acid (0.03", w,'v) and either 50 mM-physostigmine hydrochloride (Eserine) with 5 pl 14C choline (methyl I4C choF R A C T I O N NUMBER
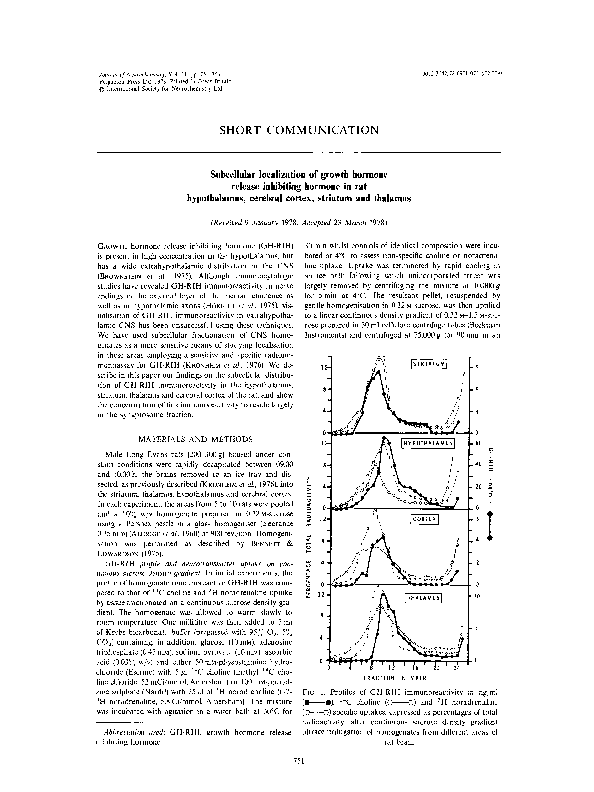
line chloride. 52 mCi/mmol, Amersham) or 100 mM-phenelzine sulphate (Nardil) with 25 pl of 'H noradrenaline (L-7- FIG. 1. Profiles of GH-RIH immunoreactivity in ng,ml
'H noradrenaline, 5.8 Ci/mmol, Amersham). The mixture (+a),
I4C choline (-0)
and 'H noradrenaline
was incubated with agitation in a water bath at 30°C for (@--O) specific uptakes. expressed as percentages of total
radioactivity. after continuous sucrose density gradient
Abhrei*iation used: GH-RIH, growth hormone release ultracentrifugation of homogenates from different areas of
inhibiting hormone.
rat brain.
751
�Short communication
752
PERCENTAGE
OF
zyxwvutsrqpo
GRADIENT
CONCEN? RATION
zyxwvutsr
zyxwvutsrqp
zyxwvutsrq
zyxwvutsrqp
zyxwvutsrqp
FIG. 2. Distribution of protein, succinic cytochrome c reductase activity (SCCR), occluded lactate dehydrogenase activity (LDH) and GH-RIH immunoreactivity on discontinuous sucrose density gradients
from rat cortex, hypothalamus, striaturn and thalamus. See text for details. MY represents 0.32 ~ - 0 . 8M
interface (myelin), S is 0.8 M-1.2 k interface (synaptosomes) and M represents the pellet (mitochondria).
SW 25.1 rotor (Spinco). The tubes were punctured and I ml
fractions of the gradient collected. 100 pl was solubilised
in 500 pl Soluene (Packard Instruments), suspended in
8oOpI Instagel (Packard Instruments) and counted in a
Packard Automatic Liquid Scintillation Counter. The
remainder was snap frozen and stored until assayed in
dilution for GH-RIH. The same homogenisation and density gradient centrifugation procedure was followed on
three occasions per area, without prior incubation, to
ensure that the GH-RIH profile was unchanged.
Production of a purified synaptosomal fraction. Following
homogenisation of brain tissue, as described above, a purified synaptosome fraction was prepared by the method of
(1962) as modified by BENNE-IT&
GRAY& WHITTAKER
EDWARDSON
(1975). The purity of the final preparation was
assessed for contamination by microsomes using D-ghCOSe6-phosphate phosphohydrolase (glucose-6-phosphatase)
et al., 1967). and mitoEC 3.1.3.9, activity (DE LAMIRANDE
chondria (using succinic-cytochrome c reductase-complex
11-111 activity) (TISDALE,
1967). Occluded lactate dehydrogenase @-lactate: NAD oxidoreductase) EC 1.1.1.27 activity was used as a marker for intact synaptosomes
(MARCHBANKS,
1967). Protein was measured by the method
of L o m v et al. (1951). Samples of all fractions were made
up to 2M-acetic acid, well mixed, centrifuged and the
supernatant freeze dried for GH-RIH assay.
G H - R l H radioimmunoassay. The radioimmunoassay for
GH-RIH was performed as previously described (KRONHEIM et al., 1976) using rabbit-anti GH-RIH haemocyanin
(1 : 125,000 final concentration), L Z 5 1 - T ~ 1 - G H - R Itracer
H
(750 pCi/pg). synthetic cyclic GH-RIH standards in serial
dilution (Ayerst AY-24910) and dextran coated charcoal,
in the presence of inactivated horse serum, to separate anti-
body bound tracer from free. The assay is sensitive to 7 ppi
assay tube and has been demonstrated to be specific for
GH-RIH.
RESULTS
Continuous sucrose density gradient ultracentrifugation
of homogenised hypothalamus, thalamus, striatum and cerebral cortex revealed a peak of GH-RIH immunoreactivity,
coinciding with the peaks of specific 14C choline and 'H
noradrenaline uptake (Fig. I). The GH-RIH peak retained
the same sedimentation characteristics when studied without preceding incubation. Another peak of GH-RIH immunoreactivity in high concentration occurred in the
0.32 M area of the gradient coinciding with unincorporated
14C choline or 3H noradrenaline.
The purified synaptosomal fraction obtained following
differential centrifugation and ultracentrifugation on a discontinuous sucrose gradient contained &60% of the
GH-RIH immunoreactivity recovered in all areas studied.
This immunoreactivity coincided with the major concentration of occluded lactic dehydrogenase activity and gradient protein as well as mitochondria1 marker enzyme (succinic cytochrome C reductase) activity (Fig. 2). Electron
microscopy performed on glutaraldehyde fixed samples of
all fractions confirmed the major concentration of intact
synaptosomes to coincide with GH-RIH immunoreactivity.
DISCUSSION
GH-RIH immunoreactivity has been shown (KRONHEIM
et al., 1976; KOBAYASHI
et al., 1977; PALKOVITS
et al., 1976)
to have a wide distribution in the CNS with the highest
�zyxwvut
zyxwvutsr
zyxwvut
zyxwvutsrqponm
zyxwvutsrq
zyxwvu
zyxwvutsrqponm
zyxwvutsrqp
zyxwvut
Short communication
753
concentrations in the hypothalamus and septum and Energy Board, the University of Cape Town Staff Research
preoptic areas hut with considerable levels in extrahypo- Fund and Nellie Atkinson Bequest. and the International
thalamic CNS. namely thalamus, striatum, cerebral cortex, Atomic Energy Agency (Contract No. 1806/RB). We thank
brain stem and spinal cord. Immunohistochemistry has the University of Cape Town Merrin Bequest for support
revealed the presence of GH-RIH immunoreactivity in of G.W.B. during his stay at the University of Cape Town.
hypothalamus alone, where it is located in nerve endings
in the median eminence and axons in the hypothalamic Isotope and Immunoassay Laboratory.
M. BtRELOWlTZ
A. HUDSON
nuclei, although some workers have been successful in Department of Medicine.
demonstrating GH-RIH containing perikarya in the peri- University of Cape Town Medical School, B. PIMSTONE'.'
S. KRONHEIM
ventricular areas of the anterior hypothalamus (HOKFELT Republic of South Africa
G. W. BENNETT'
et a/.. 1975). The reason for the more diffuse localisation and
of immunoreactive GH-RIH as measured by immunoassay Department of Physiology,
probably lies in the much greater sensitivity of the tech- St. George's Hospital Medical School,
nique: GH-RIH has been demonstrated to have an inhibi- London SW17 OPT.
tory effect on nerve activity following microiontophoretic England
application (RENAUDet a/., 1975) and has been shown to
decrease seizure duration in rats given strychnine, increasREFERENCES
ing the LD50 of the toxin (BROWN
& VALE,1975). Behavioural effects have been demonstrated following direct
ALDRIDGEW. M., EMERYR. C. & STREET
B. W. (1960)
GH-RIH application to amygdala ( R ~ Z E Ket al., 1977a),
Biochem. J. 17, 326-327.
neostriatal areas (REZEK et a!., 1977b) or cerebral cortex
BENNETT
G. W. & EDWARDSON
J. A. (1975) J. Endocririol.
(REZEKet a/., 1976) of the rat.
65, 33-44.
The wide CNS distribution of GH-RIH, together with
BENNETT
G. W. & EDWARDSONJ. A. (1977) J. Endrocripzoi.
its effects on nerve activity and behaviour, place it in the
73, 27-28P.
same category as other neuropeptides with similar properBROWN M. & VALE W. (1975) Endocrinology %,
ties. Many of these peptides including thyrotrophin-releas1333-1 336.
ing hormone, TRH (BENNETT
& EDWARDSON,
1977), suhBROWNSTEINM., ARIMURAA,, SATOH.. SCHALLY
A. V.
stance P (DUFFY
e t al., 1975), neurotensin (UHL& SNYDER,
& KIZERJ. S. (1975) Endocrinology %, 1456- 1461.
1976, and vasoactive intestinal polypeptide (GIACHETTI
et
DE LAMIRANDE
G., MORAlS R. & BLACKSTEIN
M. (1967)
al., 1977) have been shown to be concentrated in nerve
Archs Biochem. Biophys. 118, 347-351.
endings and have been postulated to comprise a new group DUFFYM. J., MULHALL
D. & POWELL
D. (1975) J . Neuroof peptidergic neurotransmitters. EPELBAUMet al. (1977)
chern. 25, 305-307.
have previously demonstrated a suhcellular localisation of
EPELBAUM
J., BRAZEAU
P., TSANGD., BRAWER
J. & MARGH-RIH in the synaptosome fraction of hypothalamus,
TIN J. B. (1977) Brain Res. 126, 309-323.
preoptic area and amygdala, areas of importance in regulaGIACHETTI
A,, SAID S. I., REYNOLDS R. C. & K O N I GF.
~ SC.
tion of pituitary growth hormone secretion (EPELBAUM
et
(1977) Proc. Natn. Acad. Sci., U.S.A. 74, 3424-3428.
al., 1977). We confirm the hypothalamic localisation of GRAYE. G. & WHITTAKER
V. P. (1962) J. Anal., Lond.
GH-RIH in nerve endings but also find a similar subcellu98, 79-87.
lar distribution in striatum, thalamus. and cerebral cortex.
HOKFELT
T., EFENDIC
S., HELLERSTROM
C., JOHANSSON 0..
suggesting a more diffuse role for the peptide.
LUFTR . & ARIMURAA. (1975) Acta endocririol. 80, Suppl.
That not all the GH-RIH recovered from the gradient
200, 5-41.
is present in our synaptosome preparation is shown by the
KOBAYASHI
R. M., BROWNM. & VALEW. (1977) Brain
presence of non-sedimented GH-RIH immunoreactivity
Res. 126, 548-588.
coinciding with free I4C choline and 'H noradrenaline.
KRONHEIM
S., BERELOWITZ
M. & PIMSTONE
B. L. (1976)
Whether this is a function of synaptosome rupture during
Clin. Endocrinol. 5, 619-630.
homogenisation or represents GH-RIH in axons or periLOWRY
H. O., ROSEBROUGHN. J., FARRA. L. & RANDALL
karya released during homogenisation is unclear.
R . J. (1951) J. hiol. Chem. 53, 265-275.
The localisation of GH-RIH in nerve endings in extraMARCHBANKS
R . M. (1967) Biochem. J. 104, 148-157.
hypothalamic CNS suggests a neurotransmitter role for the
PALKOVITS
M., BROWNSTEIN
M. J., ARIMURA
A,, SATOH..
peptide whilst its presence in synaptosomes of the median
SCHALLY
A. V. & KlzER J. S. (1976) Brain Res. 109,
eminence is consistent with a hypophysiotrophic role on
43w34.
growth hormone secretion. Further studies on the mechanRENAUDL. P., MARTIN
J. B. & BRAZEAU
P. (1975) Nature
isms controlling release of synaptosomal GH-RIH in
255, 233-235.
various CNS sites may elucidate differential physiologic
REZEK M., HAVLICEK
V., HUGHES
K. R . & FRIESEN
H.
functions.
(1976) Pharmacol. Biochem. Behac. 5, 73-77.
REZEK M., HAVLICEK
V., HUGHESK. R. & FRIESEN
H.
Acknowledye~nentsFinancialsupport was obtained from
(1977a) Neuropharmacology 16, 157-1 62.
the South African Medical Research Council and Atomic REZEKM., HAVLICEK
v., LEYBlN L., PINSKY c., KROEGER
E. A., HUGHES
K. R. & FRIESEN
H. (19776) Carl. J. Physiol. Pharmac. 55, 234-242.
TISDALE
H. D. (1967) in Methods in Enzymology (ESTATo whom correspondence should be sent.
BROOK R. W. & PULLMAN
M. E., eds.) Vol. X, pp.
'To whom reprint requests should be sent, at Depart213-215. Academic Press, New York.
ment of Medicine, University of Capetown Medical
UHLG. R . & SNYDERS. H. (1976) L f e Sci. 19, 1827--1832.
School. Observatory 7925, Republic of South Africa.
'
�
 Shirley Kronheim
Shirley Kronheim