

Structural Characterization of Polysaccharides from Noni (Morinda citrifolia L.) Juice and Their Preventive Effect on Oxidative Stress Activity
 and
and
Abstract
:1. Introduction
2. Results
2.1. Purification of NJSP
2.2. Characterization of NJSPd−1
2.2.1. Analysis of Molecular Weight and Monosaccharide Composition of Noni Juice Polysaccharide
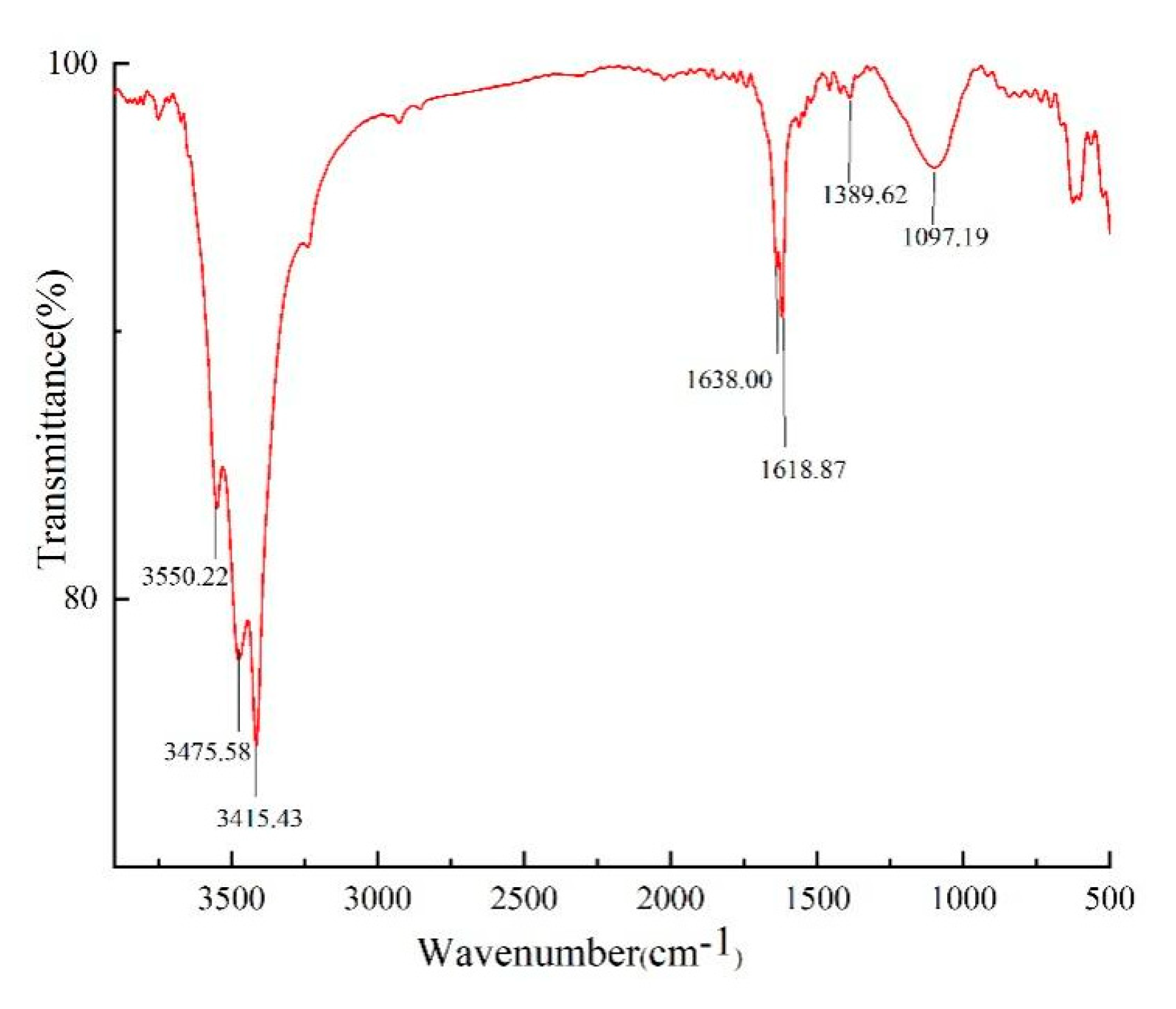
2.2.2. FT-IR Spectrum Analysis of Noni Juice Polysaccharide
2.2.3. NMR Spectrum Analysis
2.3. Effect of Different Concentrations of NJSPd−1 on the Viability of HepG2 Cells
2.4. Effects of NJSPd−1 on Oxidative Stress in HepG2 Cells
2.5. Western Blot Analysis
2.6. qPCR Analysis
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Extraction
4.3. Purification of Crude Polysaccharide
4.4. Characterization of NJSPd−1
4.4.1. Homogeneity and Molecular Weight
4.4.2. Analysis of Monosaccharide Composition
4.4.3. FT-IR Spectrometric Analysis
4.4.4. NMR Spectrometric Analysis
4.5. Cell Treatment and Grouping
4.6. Cell Activity Assay
4.7. ROS Detection
4.8. Western Blot Method
4.9. qPCR Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dilworth, L.; Facey, A.; Omoruyi, F. Diabetes mellitus and its metabolic complications: The role of adipose tissues. Int. J. Mol. Sci. 2021, 22, 7644. [Google Scholar] [CrossRef] [PubMed]
- International Diabetes Federation. IDF Diabetes Atlas (10th ed. 2021). 2022. Available online: https://diabetesatlas.org/data/en/indicators/1/ (accessed on 25 December 2024).
- Kanwugu, O.N.; Glukhareva, T.V.; Danilova, I.G.; Kovaleva, E.G. Natural Antioxidants in diabetes treatment and management: Prospects of astaxanthin. Crit. Rev. Food Sci. 2022, 62, 5005–5028. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.R.; Liu, Z.; Liu, G.Y.; Wang, H.J. Research progress of active ingredients of Scutellaria baicalensis in the treatment of type 2 diabetes and its complications. Biomed. Pharmacother. 2022, 148, 112690. [Google Scholar] [CrossRef]
- Xu, N.; Zhou, Y.J.; Lu, X.Y.; Chang, Y.N. Auricularia auricula–judae (Bull.) polysaccharides improve type 2 diabetes in HFD/STZ–induced mice by regulating the AKT/AMPK signaling pathways and the gut microbiota. J. Food Sci. 2021, 86, 5479–5494. [Google Scholar] [CrossRef]
- Demir, S.; Nawroth, P.P.; Herzig, S.; Ustunel, B.E. Emerging targets in type 2 diabetes and diabetic complications. Adv. Sci. 2021, 8, 2100275. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Farrokhi, F.R.; Butler, A.E.; Sahebkar, A. Insulin resistance: Review of the underlying molecular mechanisms. J. Cell. Physiol. 2019, 234, 8152–8161. [Google Scholar] [CrossRef]
- Boden, G.; Homko, C.; Barrero, A.C.; Stein, T.P.; Chen, X.H.; Cheung, P. Excessive caloric intake acutely causes oxidative stress, GLUT4 carbonylation, and insulin resistance in healthy men. Sci. Transl. Med. 2015, 7, 304re7. [Google Scholar] [CrossRef]
- Hu, Y.J.; Yan, Z.P.; He, Y.; Li, Y.; Li, M.; Li, Y. Ameliorative effects of different doses of selenium against fluoride-triggered apoptosis and oxidative stress-mediated renal injury in rats through the activation of Nrf2/HO-1/NQO1 signaling pathway. Food Chem. Toxicol. 2023, 174, 113647. [Google Scholar] [CrossRef]
- Wang, K.Y.; Huang, K.X.; Li, X.Q.; Wu, H.; Wang, L.; Bai, F.Y. Kelp nanocellulose combined with fucoxanthin achieves lipid-lowering function by reducing oxidative stress with activation of Nrf2/HO-1/NQO1 pathway. Food Chem. 2025, 464, 141588. [Google Scholar] [CrossRef]
- Yang, X.B.; Lin, C.R.; Cai, S.; Li, W.Z.; Tang, J.; Wu, X.Y. Therapeutic effects of noni fruit water extract and polysaccharide on oxidative stress and inflammation in mice under high-fat diet. Food Funct. 2020, 11, 1133–1145. [Google Scholar] [CrossRef]
- Algenstaedt, P.; Stumpenhagen, A.; Westendorf, J. The effect of Morinda citrifolia L. fruit juice on the blood sugar level and other serum parameters in patients with diabetes type 2. Evid.-Based Complement. Altern. Med. 2018, 2018, 3565427. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.Q.; Dai, G.H.; Jiang, S.P.; Lu, Y.; Tong, Y.P.; Wang, P. Effect of two kinds of fermented food on blood glucose and lipids in induced diabetic mice. Chin. J. Appl. Physiol. 2018, 34, 130–133. [Google Scholar] [CrossRef]
- Shoeb, A.; Alwar, M.; Shenoy, P.J.; Gokul, P. Effect of Morinda citrifolia (Noni) fruit juice on high fat diet induced dyslipidemia in rats. J. Clin. Diagn. Res. 2016, 10, FF06–FF10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Meng, J.; Li, X.J.; Tang, X.; Ma, S.H.; Lv, Y.P. Noni (Morinda citrifolia L.) wine prevents the oxidative stress and obesity in mice induced by high-fat diet. J. Food Biochem. 2020, 44, e13460. [Google Scholar] [CrossRef]
- Ahmad, A.; Alkharfy, K.M.; Wani, T.A.; Raish, M. Application of Box-Behnken design for ultrasonic-assisted extraction of polysaccharides from Paeonia emodi. Int. J. Biol. Macromol. 2015, 72, 990–997. [Google Scholar] [CrossRef]
- Jin, M.Y.; Wang, Y.X.; Yang, X.B.; Yin, H.; Nie, S.P.; Wu, X.Y. Structure characterization of a polysaccharide extracted from noni (Morinda citrifolia L.) and its protective effect against DSS-induced bowel disease in mice. Food Hydrocolloids 2019, 90, 189–197. [Google Scholar] [CrossRef]
- Sherahi, M.H.; Fathi, M.; Zhandari, F.; Hashemi, S.M.B.; Rashidi, A. Structural characterization and physicochemical properties of Descurainia sophia seed gum. Food Hydrocolloids 2017, 66, 82–89. [Google Scholar] [CrossRef]
- Xu, Y.Q.; Gao, Y.K.; Liu, F.; Niu, X.J.; Wang, L.B.; Li, X.G. Sulfated modification of the polysaccharides from blackcurrant and their antioxidant and α-amylase inhibitory activities. Int. J. Biol. Macromol. 2018, 109, 1344–1354. [Google Scholar] [CrossRef]
- Beigi, M.; Jahanbin, K. A water-soluble polysaccharide from the roots of Eremurus spectabilis M. B. subsp. spectabilis: Extraction, purification and structural features. Int. J. Biol. Macromol. 2019, 128, 648–654. [Google Scholar] [CrossRef]
- Choi, J.W.; Synytsya, A.; Capek, P.; Bleha, R.; Pohl, R.; Park, Y.I. Structural analysis and anti-obesity effect of a pectic polysaccharide isolated from Korean mulberry fruit Oddi (Morus alba L.). Carbohydr. Polym. 2016, 146, 187–196. [Google Scholar] [CrossRef]
- Pu, X.Y.; Ma, X.L.; Liu, L.; Ren, J.; Li, H.B.; Li, X.Y. Structural characterization and antioxidant activity in vitro of polysaccharides from angelica and astragalus. Carbohydr. Polym. 2016, 137, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lei, Z.X.; Zhao, M.M.; Wu, C.Z.; Wang, L.B.; Xu, Y.Q. Microwave assisted extraction of an acidic polysaccharide from Ribes nigrum L.: Structural characteristics and biological activities. Ind. Crops Prod. 2020, 147, 112249. [Google Scholar] [CrossRef]
- Jia, X.J.; Hu, J.; He, M.X.; Zhang, Q.W.; Li, P.; Wan, J.B. α-Glucosidase inhibitory activity and structural characterization of polysaccharide fraction from Rhynchosia minima root. J. Funct. Foods. 2017, 28, 76–82. [Google Scholar] [CrossRef]
- Liu, H.L.; Jiang, Y.M.; Yang, H.S.; Yang, B. Structure characteristics of an acidic polysaccharide purified from Banana (Musa nana Lour.) pulp and its enzymatic degradation. Int. J. Biol. Macromol. 2017, 101, 299–303. [Google Scholar] [CrossRef]
- Xu, Y.; Guo, Y.; Duan, S.; Wei, H.; Liu, Y.; Wang, L.; Huo, X.; Yang, Y. Effects of ultrasound irradiation on the characterization and bioactivities of the polysaccharide from blackcurrant fruits. Ultrason. Sonochem. 2018, 49, 206–214. [Google Scholar] [CrossRef]
- Tu, W.S.; Zhu, J.H.; Bi, S.X.; Chen, D.H.; Song, L.Y.; Wang, L.S. Isolation, characterization and bioactivities of a new polysaccharide from Annona squamosa and its sulfated derivative. Carbohydr. Polym. 2016, 152, 287–296. [Google Scholar] [CrossRef]
- Liu, W.; Liu, Y.M.; Zhu, R.; Yu, J.P.; Lu, W.S.; Pan, C. Structure characterization, chemical and enzymatic degradation, and chain conformation of an acidic polysaccharide from Lycium barbarum L. Carbohydr. Polym. 2016, 147, 114–124. [Google Scholar] [CrossRef]
- Zhang, Z.L.; Kong, F.S.; Ni, H.; Mo, Z.X.; Wan, J.B.; Hua, D.F. Structural characterization, α-glucosidase inhibitory and DPPH scavenging activities of polysaccharides from guava. Carbohydr. Polym. 2016, 144, 106–114. [Google Scholar] [CrossRef]
- Lohani, M.; Majrashi, M.; Govindarajulu, M.; Patel, M.; Ramesh, S.; Bhattacharya, D.; Joshi, S.; Fadan, M.; Nadar, R.; Darien, B.; et al. Immunomodulatory actions of a Polynesian herb Noni (Morinda citrifolia) and its clinical applications. Complement. Ther. Med. 2019, 47, 102206. [Google Scholar] [CrossRef]
- Zhang, X.G.; Zhang, B.; Li, L.; Li, X.B.; Zhang, J.Q.; Chen, G.Y. Fermented Noni (Morinda citrifolia L.) fruit juice improved oxidative stress and insulin resistance under the synergistic effect of Nrf2/ARE pathway and gut flora in db/db mice and HepG2 cells. Food Funct. 2022, 13, 8254–8273. [Google Scholar] [CrossRef]
- Buettner, G.R. The pecking order of free radicals and antioxidants: Lipid peroxidation, α-tocopherol, and ascorbate. Arch. Biochem. Biophys. 1993, 300, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Zeb, A. Concept, mechanism, and applications of phenolic antioxidants in foods. J. Food Biochem. 2020, 44, e13394. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.L.; Zhao, B.; Sun, S.L.; Yu, S.F.; Wang, Y.M.; Ji, R.; Yang, Z.T.; Ma, L.; Yao, Y.; Chen, Y.; et al. High-dose vitamin C alleviates pancreatic injury via the NRF2/NQO1/HO-1 pathway in a rat model of severe acute pancreatitis. Ann. Transl. Med. 2020, 8, 852. [Google Scholar] [CrossRef]
- Thirumavalavan, M.; Lai, Y.L.; Lin, L.C.; Lee, J.F. Cellulose-based native and surface modified fruit peels for the adsorption of heavy metal ions from aqueous solution: Langmuir adsorption isotherms. J. Chem. Eng. Data 2010, 55, 1186–1192. [Google Scholar] [CrossRef]
- Ji, X.L.; Cheng, Y.Q.; Tian, J.Y.; Zhang, S.Q.; Jing, Y.S.; Shi, M.M. Structural characterization of polysaccharide from jujube (Ziziphus jujuba Mill.) fruit. Chem. Biol. Technol. Agric. 2021, 8, 54. [Google Scholar] [CrossRef]
- Li, J.; Niu, D.B.; Zhang, Y.; Zeng, X.A. Physicochemical properties, antioxidant and antiproliferative activities of polysaccharides from Morinda citrifolia L. (Noni) based on different extraction methods. Int. J. Biol. Macromol. 2020, 150, 114–121. [Google Scholar] [CrossRef]
- Wang, Z.J.; Luo, D.H. Antioxidant activities of different fractions of polysaccharide purified from Gynostemma pentaphyllum makino. Carbohydr. Polym. 2007, 68, 54–58. [Google Scholar] [CrossRef]
- Ji, X.L.; Guo, J.H.; Ding, D.Q.; Gao, J.; Hao, L.R.; Guo, X.D.; Liu, Y.Q. Structural characterization and antioxidant activity of a novel high-molecular-weight polysaccharide from Ziziphus jujuba cv. Muzao. J. Food Meas. Charact. 2022, 16, 2191–2200. [Google Scholar] [CrossRef]
- Tian, P.F.; Zhou, D.J.; Ji, C.; Niu, C.Y.; Chen, Y.; Chen, Y. Characterization and anticoagulant activity of a fucosylated chondroitin sulfate from the sea cucumber Acaudina leucoprocta. Process Biochem. 2024, 147, 130–136. [Google Scholar] [CrossRef]
- Xie, W.J.; Liu, M.; Zhang, X.; Zhang, Y.G.; Jian, Z.H.; Xiong, X.X. Astaxanthin suppresses LPS-induced myocardial apoptosis by regulating PTP1B/JNK pathway in vitro. Int. Immunopharmacol. 2024, 127, 111395. [Google Scholar] [CrossRef]
- Gu, L.; Chi, M.; Wang, Z.; Fu, Y.; Li, N.; Niu, Y.; Yu, B.; Lin, J.; Li, C.; Zhao, G. Hydroxytyrosol downregulates inflammatory responses via Nrf2/HO-1 axis during fungal keratitis and exerts antifungal effects. Int. Immunopharmacol. 2025, 149, 114202. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yang, J.; Fang, Y.; Lou, C.; Yu, H.; Li, Y.; Lv, J.; Chen, H.; Cai, L.; Zheng, W. Vinpocetine protects against osteoarthritis by inhibiting ferroptosis and extracellular matrix degradation via activation of the Nrf2/GPX4 pathway. Phytomedicine 2024, 135, 156115. [Google Scholar] [CrossRef]
- Xu, L.; Wang, Y.; Chen, Q.; Zhu, X.; Hong, J. Propofol modulates Nrf2/NLRP3 signaling to ameliorate cigarette smoke-induced damage in human bronchial epithelial cells. Tissue Cell 2024, 88, 102341. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | ManA | GluA | GalA | Man | Glu | Gal | Rha | Fuc | Rib | Xyl | Ara |
|---|---|---|---|---|---|---|---|---|---|---|---|
| NJSPd−1 | 0.49 | 0.88 | 28.79 | 0.18 | 0.64 | 20.35 | 19.80 | 3.50 | 0.78 | 5.70 | 18.84 |
| Gene | Forward Primer Sequence (5′-3′) | Reverse Primer Sequence (5′-3′) |
|---|---|---|
| Nrf2 | CAGTCAGCGACGGAAAGAGTA | TGTGGGCAACCTGGGAGTAG |
| HO-1 | TGGCTGGCTTCCTTACCGT | ACCACCCCAACCCTGCTAT |
| IRS-1 | GGGAGGACTTGAGCTACGGT | GATGGGGTTAGAGCAGTTGGA |
| GXP1 | GCAACCAGTTTGGGCATCA | CCGTTCACCTCGCACTTCTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, B.; Wei, X.; Du, P.; Luo, H.; Hu, L.; Guan, L.; Chen, G. Structural Characterization of Polysaccharides from Noni (Morinda citrifolia L.) Juice and Their Preventive Effect on Oxidative Stress Activity. Molecules 2025, 30, 1103. https://doi.org/10.3390/molecules30051103
Zhang B, Wei X, Du P, Luo H, Hu L, Guan L, Chen G. Structural Characterization of Polysaccharides from Noni (Morinda citrifolia L.) Juice and Their Preventive Effect on Oxidative Stress Activity. Molecules. 2025; 30(5):1103. https://doi.org/10.3390/molecules30051103
Chicago/Turabian StyleZhang, Bin, Xiaoyu Wei, Peiwen Du, Huangqun Luo, Lanfang Hu, Liping Guan, and Guangying Chen. 2025. "Structural Characterization of Polysaccharides from Noni (Morinda citrifolia L.) Juice and Their Preventive Effect on Oxidative Stress Activity" Molecules 30, no. 5: 1103. https://doi.org/10.3390/molecules30051103
APA StyleZhang, B., Wei, X., Du, P., Luo, H., Hu, L., Guan, L., & Chen, G. (2025). Structural Characterization of Polysaccharides from Noni (Morinda citrifolia L.) Juice and Their Preventive Effect on Oxidative Stress Activity. Molecules, 30(5), 1103. https://doi.org/10.3390/molecules30051103