
Ligand-Independent Spontaneous Activation of Purinergic P2Y6 Receptor Under Cell Culture Soft Substrate

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid Construction
2.2. Cell Culture and Transfection
2.3. Intracellular Ca2+ Imaging Using Fura-2
2.4. Intracellular Ca2+ Imaging Using GCaMP6
2.5. Western Blotting
2.6. Statistical Analysis
3. Results
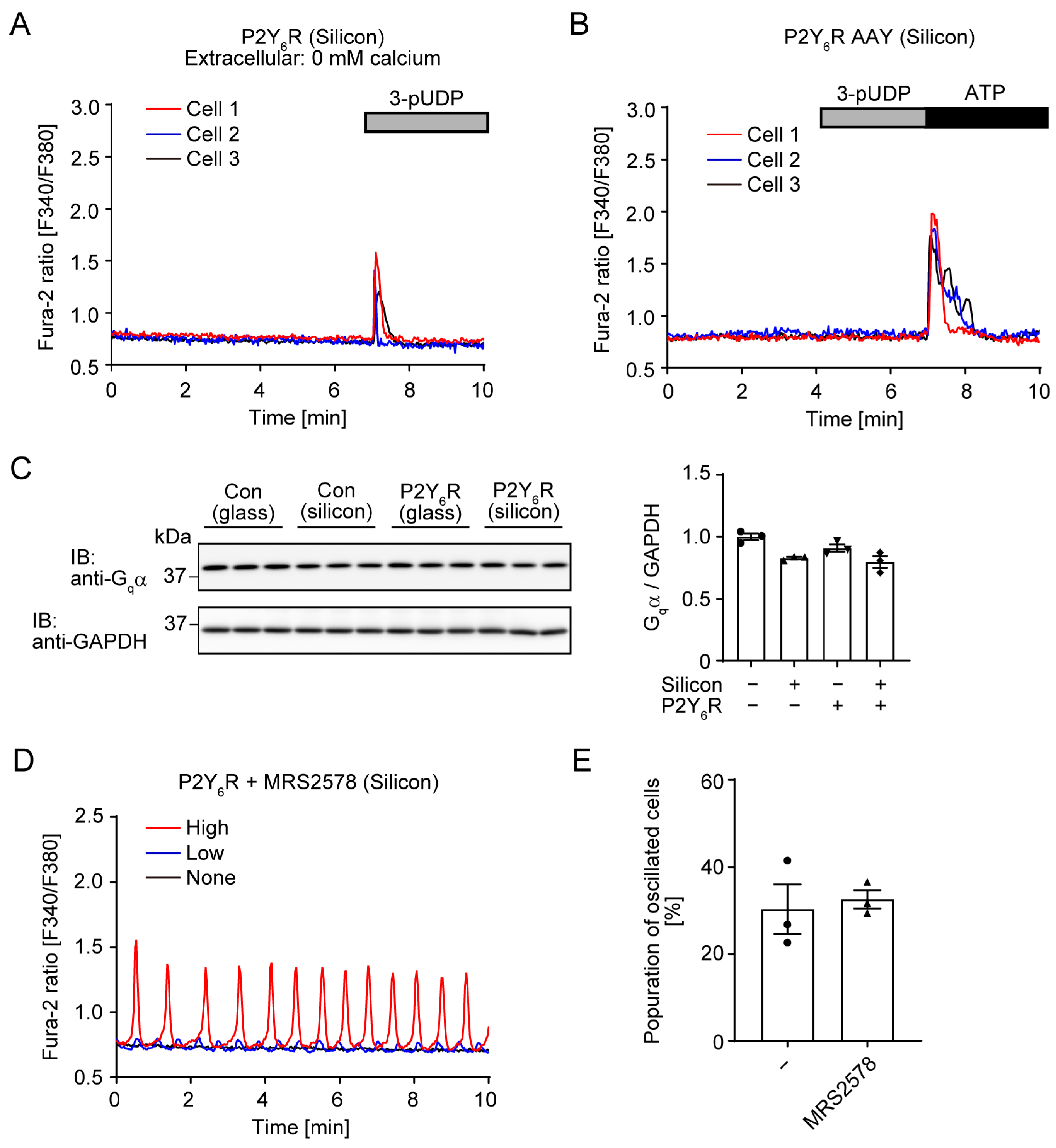
3.1. P2Y6R-Mediated Spontaneous Ca2+ Oscillation Under Silicon Chamber Culture Conditions
3.2. Effet of Extracellular Matrix Proteins on P2Y6R-Mediated Ca2+ Oscillation
3.3. G Protein-Mediated Signaling and Ca2+ Entry from Extracellular Space Is Required for Spontaneous Ca2+ Oscillation of P2Y6R
3.4. The Extracellular RGD Motif of P2Y6R Is Required for Spontaneous Ca2+ Oscillation
3.5. Ligand-Independent Ca2+ Oscillation Is Observed in Several GPCRs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fredriksson, R.; Lagerstrom, M.C.; Lundin, L.G.; Schioth, H.B. The G-protein-coupled receptors in the human genome form five main families. Phylogenetic analysis, paralogon groups, and fingerprints. Mol. Pharmacol. 2003, 63, 1256–1272. [Google Scholar] [CrossRef]
- Hauser, A.S.; Attwood, M.M.; Rask-Andersen, M.; Schioth, H.B.; Gloriam, D.E. Trends in GPCR drug discovery: New agents, targets and indications. Nat. Rev. Drug Discov. 2017, 16, 829–842. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, A.; Sunggip, C.; Oda, S.; Numaga-Tomita, T.; Tsuda, M.; Nishida, M. Purinergic P2Y receptors: Molecular diversity and implications for treatment of cardiovascular diseases. Pharmacol. Ther. 2017, 180, 113–128. [Google Scholar] [CrossRef]
- Nishiyama, K. The role of P2Y6 receptor in the pathogenesis of cardiovascular and inflammatory diseases. J. Pharmacol. Sci. 2024, 154, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Wang, W.; Li, Y.; Zhang, Q.; Ji, H.; Li, H.; Hu, Q. The role of P2Y6R in cardiovascular diseases and recent development of P2Y6R antagonists. Drug Discov. Today 2020, 25, 568–573. [Google Scholar] [CrossRef] [PubMed]
- Nishida, M.; Sato, Y.; Uemura, A.; Narita, Y.; Tozaki-Saitoh, H.; Nakaya, M.; Ide, T.; Suzuki, K.; Inoue, K.; Nagao, T.; et al. P2Y6 receptor-Galpha12/13 signalling in cardiomyocytes triggers pressure overload-induced cardiac fibrosis. EMBO J. 2008, 27, 3104–3115. [Google Scholar] [CrossRef] [PubMed]
- Daghbouche-Rubio, N.; Alvarez-Miguel, I.; Flores, V.A.; Rojo-Mencia, J.; Navedo, M.; Nieves-Citron, M.; Cidad, P.; Perez-Garcia, M.T.; Lopez-Lopez, J.R. The P2Y6 Receptor as a Potential Keystone in Essential Hypertension. Function 2024, 5, zqae045. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, A.; Sunggip, C.; Tozaki-Saitoh, H.; Shimauchi, T.; Numaga-Tomita, T.; Hirano, K.; Ide, T.; Boeynaems, J.M.; Kurose, H.; Tsuda, M.; et al. Purinergic P2Y6 receptors heterodimerize with angiotensin AT1 receptors to promote angiotensin II-induced hypertension. Sci. Signal 2016, 9, ra7. [Google Scholar] [CrossRef] [PubMed]
- Stachon, P.; Peikert, A.; Michel, N.A.; Hergeth, S.; Marchini, T.; Wolf, D.; Dufner, B.; Hoppe, N.; Ayata, C.K.; Grimm, M.; et al. P2Y6 deficiency limits vascular inflammation and atherosclerosis in mice. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2237–2245. [Google Scholar] [CrossRef] [PubMed]
- Garcia, R.A.; Yan, M.; Search, D.; Zhang, R.; Carson, N.L.; Ryan, C.S.; Smith-Monroy, C.; Zheng, J.; Chen, J.; Kong, Y.; et al. P2Y6 receptor potentiates pro-inflammatory responses in macrophages and exhibits differential roles in atherosclerotic lesion development. PLoS ONE 2014, 9, e111385. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, M.; Li, H.; Dai, C.; Yin, L.; Liu, C.; Li, Y.; Zhang, E.; Dong, X.; Ji, H.; et al. Macrophage P2Y6 receptor deletion attenuates atherosclerosis by limiting foam cell formation through phospholipase Cbeta/store-operated calcium entry/calreticulin/scavenger receptor A pathways. Eur. Heart J. 2024, 45, 268–283. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, L.; Palczewski, K. The G protein-coupled receptor rhodopsin: A historical perspective. Methods Mol. Biol. 2015, 1271, 3–18. [Google Scholar] [PubMed]
- Ohnishi, K.; Sokabe, T.; Miura, T.; Tominaga, M.; Ohta, A.; Kuhara, A. G protein-coupled receptor-based thermosensation determines temperature acclimatization of Caenorhabditis elegans. Nat. Commun. 2024, 15, 1660. [Google Scholar] [CrossRef]
- Zou, Y.; Akazawa, H.; Qin, Y.; Sano, M.; Takano, H.; Minamino, T.; Makita, N.; Iwanaga, K.; Zhu, W.; Kudoh, S.; et al. Mechanical stress activates angiotensin II type 1 receptor without the involvement of angiotensin II. Nat. Cell Biol. 2004, 6, 499–506. [Google Scholar] [CrossRef]
- Xu, J.; Mathur, J.; Vessieres, E.; Hammack, S.; Nonomura, K.; Favre, J.; Grimaud, L.; Petrus, M.; Francisco, A.; Li, J.; et al. GPR68 Senses Flow and Is Essential for Vascular Physiology. Cell 2018, 173, 762–775.e716. [Google Scholar] [CrossRef]
- Erdogmus, S.; Storch, U.; Danner, L.; Becker, J.; Winter, M.; Ziegler, N.; Wirth, A.; Offermanns, S.; Hoffmann, C.; Gudermann, T.; et al. Helix 8 is the essential structural motif of mechanosensitive GPCRs. Nat. Commun. 2019, 10, 5784. [Google Scholar] [CrossRef] [PubMed]
- Mederos y Schnitzler, M.; Storch, U.; Meibers, S.; Nurwakagari, P.; Breit, A.; Essin, K.; Gollasch, M.; Gudermann, T. Gq-coupled receptors as mechanosensors mediating myogenic vasoconstriction. EMBO J. 2008, 27, 3092–3103. [Google Scholar] [CrossRef] [PubMed]
- Chachisvilis, M.; Zhang, Y.L.; Frangos, J.A. G protein-coupled receptors sense fluid shear stress in endothelial cells. Proc. Natl. Acad. Sci. USA 2006, 103, 15463–15468. [Google Scholar] [CrossRef]
- Jung, B.; Obinata, H.; Galvani, S.; Mendelson, K.; Ding, B.S.; Skoura, A.; Kinzel, B.; Brinkmann, V.; Rafii, S.; Evans, T.; et al. Flow-regulated endothelial S1P receptor-1 signaling sustains vascular development. Dev. Cell 2012, 23, 600–610. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Majeed, S.; Nauli, S.M. Dopamine receptor type 5 in the primary cilia has dual chemo- and mechano-sensory roles. Hypertension 2011, 58, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.; Liu, J.; Xu, X.Z.S. Mechanosensitive GPCRs and ion channels in shear stress sensing. Curr. Opin. Cell Biol. 2023, 84, 102216. [Google Scholar] [CrossRef] [PubMed]
- Quitterer, U.; Fu, X.; Pohl, A.; Bayoumy, K.M.; Langer, A.; AbdAlla, S. Beta-Arrestin1 Prevents Preeclampsia by Downregulation of Mechanosensitive AT1-B2 Receptor Heteromers. Cell 2019, 176, 318–333.e319. [Google Scholar] [CrossRef] [PubMed]
- Janetzko, J.; Kise, R.; Barsi-Rhyne, B.; Siepe, D.H.; Heydenreich, F.M.; Kawakami, K.; Masureel, M.; Maeda, S.; Garcia, K.C.; von Zastrow, M.; et al. Membrane phosphoinositides regulate GPCR-beta-arrestin complex assembly and dynamics. Cell 2022, 185, 4560–4573.e4519. [Google Scholar] [CrossRef]
- Sauve, R.; Diarra, A.; Chahine, M.; Simoneau, C.; Morier, N.; Roy, G. Ca2+ oscillations induced by histamine H1 receptor stimulation in HeLa cells: Fura-2 and patch clamp analysis. Cell Calcium 1991, 12, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Thorn, P. Ca2+ influx during agonist and Ins(2,4,5)P3-evoked Ca2+ oscillations in HeLa epithelial cells. J. Physiol. 1995, 482 Pt 2, 275–281. [Google Scholar] [CrossRef]
- Chen, T.W.; Wardill, T.J.; Sun, Y.; Pulver, S.R.; Renninger, S.L.; Baohan, A.; Schreiter, E.R.; Kerr, R.A.; Orger, M.B.; Jayaraman, V.; et al. Ultrasensitive fluorescent proteins for imaging neuronal activity. Nature 2013, 499, 295–300. [Google Scholar] [CrossRef]
- Burgess, J.K.; Gosens, R. Mechanotransduction and the extracellular matrix: Key drivers of lung pathologies and drug responsiveness. Biochem. Pharmacol. 2024, 228, 116255. [Google Scholar] [CrossRef]
- Stanton, A.E.; Tong, X.; Yang, F. Extracellular matrix type modulates mechanotransduction of stem cells. Acta Biomater. 2019, 96, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Feng, Y.; Chen, X.; Gerard, C.; Boisvert, W.A. Characterization of G protein coupling mediated by the conserved D134(3.49) of DRY motif, M241(6.34), and F251(6.44) residues on human CXCR1. FEBS Open Bio 2015, 5, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Ahn, S.; Shenoy, S.K.; Karnik, S.S.; Hunyady, L.; Luttrell, L.M.; Lefkowitz, R.J. Independent beta-arrestin 2 and G protein-mediated pathways for angiotensin II activation of extracellular signal-regulated kinases 1 and 2. Proc. Natl. Acad. Sci. USA 2003, 100, 10782–10787. [Google Scholar] [CrossRef] [PubMed]
- Corriden, R.; Insel, P.A. Basal release of ATP: An autocrine-paracrine mechanism for cell regulation. Sci. Signal 2010, 3, re1. [Google Scholar] [CrossRef] [PubMed]
- Mamedova, L.K.; Joshi, B.V.; Gao, Z.G.; von Kugelgen, I.; Jacobson, K.A. Diisothiocyanate derivatives as potent, insurmountable antagonists of P2Y6 nucleotide receptors. Biochem. Pharmacol. 2004, 67, 1763–1770. [Google Scholar] [CrossRef] [PubMed]
- Ruoslahti, E. RGD and other recognition sequences for integrins. Annu. Rev. Cell Dev. Biol. 1996, 12, 697–715. [Google Scholar] [CrossRef] [PubMed]
- Greenspoon, N.; Hershkoviz, R.; Alon, R.; Varon, D.; Shenkman, B.; Marx, G.; Federman, S.; Kapustina, G.; Lider, O. Structural analysis of integrin recognition and the inhibition of integrin-mediated cell functions by novel nonpeptidic surrogates of the Arg-Gly-Asp sequence. Biochemistry 1993, 32, 1001–1008. [Google Scholar] [CrossRef] [PubMed]
- Erb, L.; Liu, J.; Ockerhausen, J.; Kong, Q.; Garrad, R.C.; Griffin, K.; Neal, C.; Krugh, B.; Santiago-Perez, L.I.; Gonzalez, F.A.; et al. An RGD sequence in the P2Y2 receptor interacts with αVβ3 integrins and is required for Go-mediated signal transduction. J. Cell Biol. 2001, 153, 491–501. [Google Scholar] [CrossRef]
- Lazarowski, E.R.; Sesma, J.I.; Seminario-Vidal, L.; Kreda, S.M. Molecular mechanisms of purine and pyrimidine nucleotide release. Adv. Pharmacol. 2011, 61, 221–261. [Google Scholar] [PubMed]
- Taruno, A. ATP Release Channels. Int. J. Mol. Sci. 2018, 19, 808. [Google Scholar] [CrossRef]
- Riquelme, M.A.; Cardenas, E.R.; Xu, H.; Jiang, J.X. The Role of Connexin Channels in the Response of Mechanical Loading and Unloading of Bone. Int. J. Mol. Sci. 2020, 21, 1146. [Google Scholar] [CrossRef] [PubMed]
- Workman, A.D.; Carey, R.M.; Chen, B.; Saunders, C.J.; Marambaud, P.; Mitchell, C.H.; Tordoff, M.G.; Lee, R.J.; Cohen, N.A. CALHM1-Mediated ATP Release and Ciliary Beat Frequency Modulation in Nasal Epithelial Cells. Sci. Rep. 2017, 7, 6687. [Google Scholar] [CrossRef]
- Liu, H.T.; Toychiev, A.H.; Takahashi, N.; Sabirov, R.Z.; Okada, Y. Maxi-anion channel as a candidate pathway for osmosensitive ATP release from mouse astrocytes in primary culture. Cell Res. 2008, 18, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Singer, G.; Araki, T.; Weijer, C.J. Oscillatory cAMP cell-cell signalling persists during multicellular Dictyostelium development. Commun. Biol. 2019, 2, 139. [Google Scholar] [CrossRef] [PubMed]
- Petersen, S.C.; Luo, R.; Liebscher, I.; Giera, S.; Jeong, S.J.; Mogha, A.; Ghidinelli, M.; Feltri, M.L.; Schoneberg, T.; Piao, X.; et al. The adhesion GPCR GPR126 has distinct, domain-dependent functions in Schwann cell development mediated by interaction with laminin-211. Neuron 2015, 85, 755–769. [Google Scholar] [CrossRef]
- Wang, Y.G.; Samarel, A.M.; Lipsius, S.L. Laminin binding to beta1-integrins selectively alters beta1- and beta2-adrenoceptor signalling in cat atrial myocytes. J. Physiol. 2000, 527 Pt 1, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Shin, S.; Bae, S.W. cAMP Biosensors Based on Genetically Encoded Fluorescent/Luminescent Proteins. Biosensors 2021, 11, 39. [Google Scholar] [CrossRef] [PubMed]
- Sajkowska, J.J.; Tsang, C.H.; Kozielewicz, P. Application of FRET- and BRET-based live-cell biosensors in deorphanization and ligand discovery studies on orphan G protein-coupled receptors. SLAS Discov. 2024, 29, 100174. [Google Scholar] [CrossRef] [PubMed]
- De Belly, H.; Paluch, E.K.; Chalut, K.J. Interplay between mechanics and signalling in regulating cell fate. Nat. Rev. Mol. Cell Biol. 2022, 23, 465–480. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Zhou, Q.; Liu, C.; Zhang, Y.; Xie, M.; Qiao, W.; Dong, N. Substrate stiffness regulates differentiation of induced pluripotent stem cells into heart valve endothelial cells. Acta Biomater. 2022, 143, 115–126. [Google Scholar] [CrossRef]
- Wei, J.; Yao, J.; Yan, M.; Xie, Y.; Liu, P.; Mao, Y.; Li, X. The role of matrix stiffness in cancer stromal cell fate and targeting therapeutic strategies. Acta Biomater. 2022, 150, 34–47. [Google Scholar] [CrossRef]
- Atcha, H.; Jairaman, A.; Holt, J.R.; Meli, V.S.; Nagalla, R.R.; Veerasubramanian, P.K.; Brumm, K.T.; Lim, H.E.; Othy, S.; Cahalan, M.D.; et al. Mechanically activated ion channel Piezo1 modulates macrophage polarization and stiffness sensing. Nat. Commun. 2021, 12, 3256. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, M.; Mamistvalov, R.; Sprinzak, D.; Vollmar, A.M.; Zahler, S. Matrix stiffness regulates Notch signaling activity in endothelial cells. J. Cell Sci. 2023, 136, jcs260442. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.J.; Sun, J.; Lu, S.; Zhang, J.; Wang, Y. The regulation of beta-adrenergic receptor-mediated PKA activation by substrate stiffness via microtubule dynamics in human MSCs. Biomaterials 2014, 35, 8348–8356. [Google Scholar] [CrossRef] [PubMed]
- Mafi, A.; Kim, S.K.; Goddard, W.A., 3rd. The mechanism for ligand activation of the GPCR-G protein complex. Proc. Natl. Acad. Sci. USA 2022, 119, e2110085119. [Google Scholar] [CrossRef] [PubMed]
- Cullum, S.A.; Platt, S.; Dale, N.; Isaac, O.C.; Wragg, E.S.; Soave, M.; Veprintsev, D.B.; Woolard, J.; Kilpatrick, L.E.; Hill, S.J. Mechano-sensitivity of beta2-adrenoceptors enhances constitutive activation of cAMP generation that is inhibited by inverse agonists. Commun. Biol. 2024, 7, 417. [Google Scholar] [CrossRef] [PubMed]
- Seifert, R.; Wenzel-Seifert, K. Constitutive activity of G-protein-coupled receptors: Cause of disease and common property of wild-type receptors. Naunyn Schmiedebergs Arch. Pharmacol. 2002, 366, 381–416. [Google Scholar] [CrossRef]
- Yu, R.; Cui, Z.; Li, M.; Yang, Y.; Zhong, J. Dimer-dependent intrinsic/basal activity of the class B G protein-coupled receptor PAC1 promotes cellular anti-apoptotic activity through Wnt/beta-catenin pathways that are associated with dimer endocytosis. PLoS ONE 2014, 9, e113913. [Google Scholar] [CrossRef]
- Stoy, H.; Gurevich, V.V. How genetic errors in GPCRs affect their function: Possible therapeutic strategies. Genes. Dis. 2015, 2, 108–132. [Google Scholar] [CrossRef] [PubMed]
- Vizurraga, A.; Adhikari, R.; Yeung, J.; Yu, M.; Tall, G.G. Mechanisms of adhesion G protein-coupled receptor activation. J. Biol. Chem. 2020, 295, 14065–14083. [Google Scholar] [CrossRef]
- Uhlen, P.; Fritz, N. Biochemistry of calcium oscillations. Biochem. Biophys. Res. Commun. 2010, 396, 28–32. [Google Scholar] [CrossRef]
- Dupont, G.; Combettes, L.; Bird, G.S.; Putney, J.W. Calcium oscillations. Cold Spring Harb. Perspect. Biol. 2011, 3, a004226. [Google Scholar] [CrossRef] [PubMed]
- Abramowitz, J.; Yildirim, E.; Birnbaumer, L. The TRPC Family of Ion Channels: Relation to the TRP Superfamily and Role in Receptor- and Store-Operated Calcium Entry. In TRP Ion Channel Function in Sensory Transduction and Cellular Signaling Cascades; Liedtke, W.B., Heller, S., Eds.; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Diarra, A.; Wang, R.; Garneau, L.; Gallo-Payet, N.; Sauve, R. Histamine-evoked Ca2+ oscillations in HeLa cells are sensitive to methylxanthines but insensitive to ryanodine. Pflugers Arch. 1994, 426, 129–138. [Google Scholar] [CrossRef]
- Lin, G.C.; Rurangirwa, J.K.; Koval, M.; Steinberg, T.H. Gap junctional communication modulates agonist-induced calcium oscillations in transfected HeLa cells. J. Cell Sci. 2004, 117, 881–887. [Google Scholar] [CrossRef]
- Hu, W.E.; Zhang, X.; Guo, Q.F.; Yang, J.W.; Yang, Y.; Wei, S.C.; Su, X.D. HeLa-CCL2 cell heterogeneity studied by single-cell DNA and RNA sequencing. PLoS ONE 2019, 14, e0225466. [Google Scholar] [CrossRef] [PubMed]
- Fukuoka, M.; Kang, W.; Horiike, S.; Yamada, M.; Miyado, K. Calcium oscillations and mitochondrial enzymes in stem cells. Regen. Ther. 2024, 26, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Gaetani, R.; Zizzi, E.A.; Deriu, M.A.; Morbiducci, U.; Pesce, M.; Messina, E. When Stiffness Matters: Mechanosensing in Heart Development and Disease. Front. Cell Dev. Biol. 2020, 8, 334. [Google Scholar] [CrossRef] [PubMed]
- Bloksgaard, M.; Lindsey, M.; Martinez-Lemus, L.A. Extracellular matrix in cardiovascular pathophysiology. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H1687–H1690. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, K.; Nishimura, A.; Sunggip, C.; Ito, T.; Nishiyama, K.; Kato, Y.; Tanaka, T.; Tozaki-Saitoh, H.; Tsuda, M.; Nishida, M. Modulation of P2Y6R expression exacerbates pressure overload-induced cardiac remodeling in mice. Sci. Rep. 2020, 10, 13926. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.H.; Trinh, T.N. P2 Receptors in Cardiac Myocyte Pathophysiology and Mechanotransduction. Int. J. Mol. Sci. 2020, 22, 251. [Google Scholar] [CrossRef]
- Kim, J.C.; Woo, S.H. Shear stress induces a longitudinal Ca2+ wave via autocrine activation of P2Y1 purinergic signalling in rat atrial myocytes. J. Physiol. 2015, 593, 5091–5109. [Google Scholar] [CrossRef]
- Cho, J.M.; Park, S.K.; Kwon, O.S.; Taylor La Salle, D.; Cerbie, J.; Fermoyle, C.C.; Morgan, D.; Nelson, A.; Bledsoe, A.; Bharath, L.P.; et al. Activating P2Y1 receptors improves function in arteries with repressed autophagy. Cardiovasc. Res. 2023, 119, 252–267. [Google Scholar] [CrossRef]
- Placet, M.; Arguin, G.; Molle, C.M.; Babeu, J.P.; Jones, C.; Carrier, J.C.; Robaye, B.; Geha, S.; Boudreau, F.; Gendron, F.P. The G protein-coupled P2Y6 receptor promotes colorectal cancer tumorigenesis by inhibiting apoptosis. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 1539–1551. [Google Scholar] [CrossRef]
- Ishihara, S.; Haga, H. Matrix Stiffness Contributes to Cancer Progression by Regulating Transcription Factors. Cancers 2022, 14, 1049. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nishimura, A.; Nishiyama, K.; Ito, T.; Mi, X.; Kato, Y.; Inoue, A.; Aoki, J.; Nishida, M. Ligand-Independent Spontaneous Activation of Purinergic P2Y6 Receptor Under Cell Culture Soft Substrate. Cells 2025, 14, 216. https://doi.org/10.3390/cells14030216
Nishimura A, Nishiyama K, Ito T, Mi X, Kato Y, Inoue A, Aoki J, Nishida M. Ligand-Independent Spontaneous Activation of Purinergic P2Y6 Receptor Under Cell Culture Soft Substrate. Cells. 2025; 14(3):216. https://doi.org/10.3390/cells14030216
Chicago/Turabian StyleNishimura, Akiyuki, Kazuhiro Nishiyama, Tomoya Ito, Xinya Mi, Yuri Kato, Asuka Inoue, Junken Aoki, and Motohiro Nishida. 2025. "Ligand-Independent Spontaneous Activation of Purinergic P2Y6 Receptor Under Cell Culture Soft Substrate" Cells 14, no. 3: 216. https://doi.org/10.3390/cells14030216
APA StyleNishimura, A., Nishiyama, K., Ito, T., Mi, X., Kato, Y., Inoue, A., Aoki, J., & Nishida, M. (2025). Ligand-Independent Spontaneous Activation of Purinergic P2Y6 Receptor Under Cell Culture Soft Substrate. Cells, 14(3), 216. https://doi.org/10.3390/cells14030216