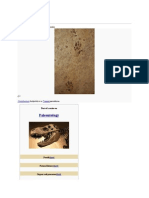
Oldest Coelacanth, From The Early Devonian of Australia: References Email Alerting Service
Oldest Coelacanth, From The Early Devonian of Australia: References Email Alerting Service
Download as pdf or txt
You might also like
- Pelvic Girdle and Fin of Tiktaalik Roseae PDFDocument7 pagesPelvic Girdle and Fin of Tiktaalik Roseae PDFSofiaJaneChavezNo ratings yet
- Week 3-6 Effect of UV On Yeast S18Document6 pagesWeek 3-6 Effect of UV On Yeast S18BlackBunny103No ratings yet
- AJP Motos - PR5 Parts List - 250Document52 pagesAJP Motos - PR5 Parts List - 250AJP MotosNo ratings yet
- Phil. Trans. R. Soc. B-2010-Ward-3333-44Document13 pagesPhil. Trans. R. Soc. B-2010-Ward-3333-44carlosart26No ratings yet
- Origins of CetaceansDocument17 pagesOrigins of CetaceansVengadesh Krishna KumarNo ratings yet
- L33 Evolution LifeDocument40 pagesL33 Evolution LifeJam Gumanoy NavarroNo ratings yet
- Fossils & EvolutionDocument11 pagesFossils & EvolutionDaniel BerryNo ratings yet
- The Ancestry of Modern Amphibia: A Review of the EvidenceFrom EverandThe Ancestry of Modern Amphibia: A Review of the EvidenceNo ratings yet
- Racicot2019 Cetacea IntroDocument6 pagesRacicot2019 Cetacea Introanon_509992240No ratings yet
- Panzeri 2024 Revisiting The Cretaceous Lungfish Atlantoceratodus Iheringi, Ameghino 1898, From The Mata Amarilla Formation, With Comments On Tooth Plate HistologyDocument26 pagesPanzeri 2024 Revisiting The Cretaceous Lungfish Atlantoceratodus Iheringi, Ameghino 1898, From The Mata Amarilla Formation, With Comments On Tooth Plate HistologyCarolina Acosta HospitalecheNo ratings yet
- App057 - Pachyrhinosaurus PerotorumDocument14 pagesApp057 - Pachyrhinosaurus PerotorumKanekoNo ratings yet
- The Earliest Herbivorous Marine ReptileDocument6 pagesThe Earliest Herbivorous Marine Reptileyou knowNo ratings yet
- Senter 2003Document9 pagesSenter 2003J.D. NobleNo ratings yet
- Biostratigraphy of The Scalenodontoides Assemblage Zone (Stormberg Group, Karoo Supergroup), South AfricaDocument10 pagesBiostratigraphy of The Scalenodontoides Assemblage Zone (Stormberg Group, Karoo Supergroup), South AfricaNeo MetsingNo ratings yet
- Dino de BrasilDocument6 pagesDino de BrasilFabiolaNo ratings yet
- Gómez, R.O. 2011. A Snake Dentary From The Upper Cretaceous of PatagoniaDocument5 pagesGómez, R.O. 2011. A Snake Dentary From The Upper Cretaceous of PatagoniaRaul Orencio GomezNo ratings yet
- Uro 2 PDFDocument5 pagesUro 2 PDFMaria octavianiNo ratings yet
- 1.5 Million Homo Erectus Jaw Sangiran 4Document14 pages1.5 Million Homo Erectus Jaw Sangiran 4Popa ConstantinNo ratings yet
- The Ancestry of Modern Amphibia: A Review of the EvidenceFrom EverandThe Ancestry of Modern Amphibia: A Review of the EvidenceNo ratings yet
- Emerging Views From Comparative Biology and GeologyDocument10 pagesEmerging Views From Comparative Biology and GeologyLuis AguilarNo ratings yet
- Chen 2014Document10 pagesChen 2014Hernan Gerley Alvarado RodriguezNo ratings yet
- Early HominisDocument9 pagesEarly Hoministaddan76No ratings yet
- The_origin_of_annelidsDocument14 pagesThe_origin_of_annelidssamsul06 rizalNo ratings yet
- Burrow Et Al 2003 - Late - D - Acanthodians - CDocument7 pagesBurrow Et Al 2003 - Late - D - Acanthodians - CJuan JoseNo ratings yet
- Ward 2015 AustralopitecinosDocument14 pagesWard 2015 Australopitecinostomascostelafre1410No ratings yet
- New Megaraptorid Dinosauria Theropoda Remains From The Lower Cretaceous Eumeralla Formation of Cape Otway Victoria AustraliaDocument20 pagesNew Megaraptorid Dinosauria Theropoda Remains From The Lower Cretaceous Eumeralla Formation of Cape Otway Victoria AustraliaJoe HinchliffeNo ratings yet
- Beetles Insecta ColeopteraDocument14 pagesBeetles Insecta Coleopteradipeshsingh563No ratings yet
- Fossil - WikipediaDocument13 pagesFossil - Wikipediaskline3No ratings yet
- tmpF53E TMPDocument7 pagestmpF53E TMPFrontiersNo ratings yet
- Hou 2014Document39 pagesHou 2014carlos.emr1991No ratings yet
- Lambert 2016Document71 pagesLambert 2016Iqbal FitriantoNo ratings yet
- A Redescription of Chaoyangia Beishanensis Aves and A Comprehensive Phylogeny of Mesozoic BirdsDocument19 pagesA Redescription of Chaoyangia Beishanensis Aves and A Comprehensive Phylogeny of Mesozoic BirdsHector LealNo ratings yet
- Fauna Amphibia GeneralDocument14 pagesFauna Amphibia GeneralCamilleLouiseJaureguiNo ratings yet
- A New Teiid Lizard From The Late Cretaceous of The Haţeg Basin, Romania and Its Phylogenetic and Palaeobiogeographical RelationshipsDocument20 pagesA New Teiid Lizard From The Late Cretaceous of The Haţeg Basin, Romania and Its Phylogenetic and Palaeobiogeographical RelationshipsJ.D. NobleNo ratings yet
- The Evolution of WhalesDocument6 pagesThe Evolution of WhalesHeather ErhayelNo ratings yet
- Letter: Diverse Transitional Giant Fleas From The Mesozoic Era of ChinaDocument4 pagesLetter: Diverse Transitional Giant Fleas From The Mesozoic Era of ChinaThiago MarinhoNo ratings yet
- Organic Evolution Notes For Grade XIDocument11 pagesOrganic Evolution Notes For Grade XIssah4155No ratings yet
- 10 1016@j Geobios 2020 07 005Document27 pages10 1016@j Geobios 2020 07 005DirtyBut WholeNo ratings yet
- The Origin of Land Vertebrates Brendan WoodDocument10 pagesThe Origin of Land Vertebrates Brendan Woodapi-253305501No ratings yet
- Enigmatic Amphibians in Mid-Cretaceous Amber Were Chameleon-Like Ballistic FeedersDocument6 pagesEnigmatic Amphibians in Mid-Cretaceous Amber Were Chameleon-Like Ballistic FeedersDirtyBut WholeNo ratings yet
- EvolutionDocument4 pagesEvolutionChaudhry AzeemNo ratings yet
- J Precamres 2007 04 009Document15 pagesJ Precamres 2007 04 009Muhammad Husnain AHNo ratings yet
- Ciclostomi FosiliDocument5 pagesCiclostomi FosiliAnca MihalcescuNo ratings yet
- GauthierDocument306 pagesGauthierDan IstrateNo ratings yet
- Ryan TaphonomyCentrosaurusOrnithischia 2001Document26 pagesRyan TaphonomyCentrosaurusOrnithischia 2001Crocodilo VasconcelosNo ratings yet
- Palopolo Et Al 2021 ZoroDocument18 pagesPalopolo Et Al 2021 ZoroGeorge ThoreauNo ratings yet
- 1-s2.0-S0960982220308289 Convergent Evolution of Swimming AdaptationsDocument10 pages1-s2.0-S0960982220308289 Convergent Evolution of Swimming AdaptationsHua Hidari YangNo ratings yet
- Journal of Anatomy - 2018 - Frey - The Remarkable Vocal Anatomy of The Koala Phascolarctos Cinereus Insights IntoDocument21 pagesJournal of Anatomy - 2018 - Frey - The Remarkable Vocal Anatomy of The Koala Phascolarctos Cinereus Insights IntoNxyz GxyzNo ratings yet
- 2009 - Evidende For Modular Evolution in A Long-Tailed Pterossaur With A Pterodactyloid SkullDocument8 pages2009 - Evidende For Modular Evolution in A Long-Tailed Pterossaur With A Pterodactyloid SkullIsaacNo ratings yet
- Palaeontology - 2003 - Waloszek - A Larval Sea Spider Arthropoda Pycnogonida From The Upper Cambrian Orsten of SwedenDocument26 pagesPalaeontology - 2003 - Waloszek - A Larval Sea Spider Arthropoda Pycnogonida From The Upper Cambrian Orsten of Swedenkian.fowlerNo ratings yet
- Yan Et Al. 2014. The Most Mysterious Beetles Jurassic Jurodidae (Insecta Coleoptera) From ChinaDocument12 pagesYan Et Al. 2014. The Most Mysterious Beetles Jurassic Jurodidae (Insecta Coleoptera) From ChinaCarlos A. JulioNo ratings yet
- Temporal Framework For The Yanliao Biota and Timing of The Origin of Crown MammalsDocument11 pagesTemporal Framework For The Yanliao Biota and Timing of The Origin of Crown MammalsCrocodilo VasconcelosNo ratings yet
- Pleo 81 625.1266 1283VFinalNautiDocument19 pagesPleo 81 625.1266 1283VFinalNautigirvanellaNo ratings yet
- A Remarkable New Cave Scorpion of The Family Pseudochactidae Gromov (Chelicerata, Scorpiones) From VietnamDocument13 pagesA Remarkable New Cave Scorpion of The Family Pseudochactidae Gromov (Chelicerata, Scorpiones) From VietnamGonzalo Ramírez GalánNo ratings yet
- Sus Dentition IdentificationDocument20 pagesSus Dentition Identificationcristian micó sanchisNo ratings yet
- A New CrossognathiformDocument14 pagesA New CrossognathiformValeria BotacheNo ratings yet
- Trace FossilDocument19 pagesTrace FossilPierre Yusalman LesmanaNo ratings yet
- Po/arornis,: The Morphology and Systematics of A Cretaceous Loon (Aves: Gaviidae) From AntarcticaDocument34 pagesPo/arornis,: The Morphology and Systematics of A Cretaceous Loon (Aves: Gaviidae) From AntarcticaAmelia KumalawatiNo ratings yet
- Garwood-Edgecombe2011 - Article - EarlyTerrestrialAnimalsEvolut & TerjemahanDocument26 pagesGarwood-Edgecombe2011 - Article - EarlyTerrestrialAnimalsEvolut & TerjemahanHumaira Isna KartikaNo ratings yet
- A. GarhiDocument9 pagesA. GarhiDaysi SuarezNo ratings yet
- Necromancy and SpiritismDocument24 pagesNecromancy and SpiritismFrancis LoboNo ratings yet
- Request For Proposal ("RFP") For Library Management SystemDocument115 pagesRequest For Proposal ("RFP") For Library Management SystemRauf NoorNo ratings yet
- Paasche Talon TG Manual EnglishDocument2 pagesPaasche Talon TG Manual Englishjhon drinkwaterNo ratings yet
- MN 63489 0013 en FPX PDFDocument544 pagesMN 63489 0013 en FPX PDFAndrey KapustaNo ratings yet
- s1166501305921 PDFDocument8 pagess1166501305921 PDFRaj RajeshNo ratings yet
- Denso ADocument1 pageDenso ADobai GaborNo ratings yet
- Catalogue: Voice Alarm Control and Indicating Equipment (VACIE)Document208 pagesCatalogue: Voice Alarm Control and Indicating Equipment (VACIE)christianxlaNo ratings yet
- Teaching ProfessionDocument19 pagesTeaching ProfessionKenneth ColoradoNo ratings yet
- Data Transfer TechniquesDocument33 pagesData Transfer TechniquesPRASHANT KUMARNo ratings yet
- US Day of The Week EffectDocument23 pagesUS Day of The Week EffectRENJiiiNo ratings yet
- Sony MEX-BT38UW, BT3800U, BT3807U, BT3850U PDFDocument50 pagesSony MEX-BT38UW, BT3800U, BT3807U, BT3850U PDFboroda2410100% (2)
- 1 X TRIGON L Plus Boiler LLH 1CT Heating Circuit 1 DHW Circuit With A CalorifierDocument4 pages1 X TRIGON L Plus Boiler LLH 1CT Heating Circuit 1 DHW Circuit With A Calorifieradyro12No ratings yet
- Yugma BalaDocument1 pageYugma BalamaabhagwatiNo ratings yet
- Using Import Filter For Siemens RelayDocument16 pagesUsing Import Filter For Siemens RelaykrlalitNo ratings yet
- Vande Bharat Express - Wikipedia PDFDocument109 pagesVande Bharat Express - Wikipedia PDFAbdulqadir DhankotNo ratings yet
- Purchase OrdersDocument9 pagesPurchase OrderslulughoshNo ratings yet
- Indian Artillery ModernisationDocument5 pagesIndian Artillery ModernisationShane Kaung MyatNo ratings yet
- Naveen Kumar Resume Format5Document2 pagesNaveen Kumar Resume Format5Satish KumarNo ratings yet
- Transformer Bunds Oil Filled Substation Transformers BS EN 61936 1 2010Document4 pagesTransformer Bunds Oil Filled Substation Transformers BS EN 61936 1 2010Agung H RosenoNo ratings yet
- Tectonism in Architecture Design and Fas PDFDocument11 pagesTectonism in Architecture Design and Fas PDFrdwhaleNo ratings yet
- Obesidade X Longividade 2018Document9 pagesObesidade X Longividade 2018Fabiana Faria LimaNo ratings yet
- Agung Sutiono Pontoh (054) - Listing Sequence SummaryDocument3 pagesAgung Sutiono Pontoh (054) - Listing Sequence SummaryFarhan. MuhammadNo ratings yet
- Research PaperDocument7 pagesResearch Paperrajbir kaurNo ratings yet
- USIBC Vacancy Communications Director 1654511038Document1 pageUSIBC Vacancy Communications Director 1654511038Vicky PanditaNo ratings yet
- Open 423014402 Pipeline Repair Procedure Final MR 050112 PDFDocument35 pagesOpen 423014402 Pipeline Repair Procedure Final MR 050112 PDFthawhanNo ratings yet
- DDP 2Document31 pagesDDP 2Anand RathiNo ratings yet
- S.N Name Base Location Activity Type Date Site Id Cell IdDocument20 pagesS.N Name Base Location Activity Type Date Site Id Cell IdAmit SharmaNo ratings yet
- Project Proposal: Raja Puniani HU SIEM, SiliguriDocument44 pagesProject Proposal: Raja Puniani HU SIEM, SiliguriRaja PunianiNo ratings yet