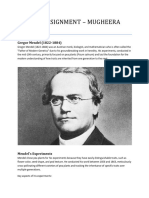
Chapter 14
Chapter 14
Download as doc, pdf, or txt
You might also like
- Ws Snorks Protein Synthesis b6 CDocument7 pagesWs Snorks Protein Synthesis b6 Capi-2331875660% (1)
- Single Gene Inheritance HandoutDocument16 pagesSingle Gene Inheritance HandoutMarel LatawanNo ratings yet
- The Teacher Will Show Scrambled Words of Recessive, Dominant, Gregor Mendel, Traits, Parents, Offspring, Heredity, Phenotype, Genotype, GenerationDocument3 pagesThe Teacher Will Show Scrambled Words of Recessive, Dominant, Gregor Mendel, Traits, Parents, Offspring, Heredity, Phenotype, Genotype, GenerationDennis Corton Tabinga100% (1)
- Inheritance ChapterDocument26 pagesInheritance Chapterainaansari2212No ratings yet
- Chapter 14 Notes 2014Document15 pagesChapter 14 Notes 2014Adeela AashiNo ratings yet
- Chapter 14 MendelDocument16 pagesChapter 14 MendelloNo ratings yet
- Law of SegregationDocument3 pagesLaw of SegregationMichael SopranoNo ratings yet
- 01 Pekan 1 - Gregor Mendel and The Laws of InheritanceDocument16 pages01 Pekan 1 - Gregor Mendel and The Laws of InheritanceEren YooNo ratings yet
- Mendel Gene Idea: and TheDocument24 pagesMendel Gene Idea: and Thekiven.samorinNo ratings yet
- Gregor MendelDocument2 pagesGregor MendelhomamunfatNo ratings yet
- MendelDocument15 pagesMendelAfaq AhmadNo ratings yet
- Mendel'S Experiments and The Laws of Probability: Openstax CollegeDocument12 pagesMendel'S Experiments and The Laws of Probability: Openstax CollegeSantiagoNo ratings yet
- GeneticsDocument32 pagesGeneticsriyanshis892No ratings yet
- Chapter 6 - Gregor Mendel and GeneticsDocument20 pagesChapter 6 - Gregor Mendel and GeneticsElizabeth100% (1)
- MendelDocument8 pagesMendelPogi BearNo ratings yet
- SBTA1301Document240 pagesSBTA1301sajithajith020No ratings yet
- Mendel's Laws of Inheritance (Notes) : CharactersDocument15 pagesMendel's Laws of Inheritance (Notes) : Characterssahoo&sahoo akashNo ratings yet
- Unit 7 HeredityDocument20 pagesUnit 7 HeredityAmriani AnrahNo ratings yet
- Hybrids: Mendel's GeneticsDocument5 pagesHybrids: Mendel's GeneticsAnonymous doCtd0IJDNNo ratings yet
- HeredityDocument10 pagesHeredity01 Jawsua Attiphat WijitNo ratings yet
- Biology Investegatory Project 4Document9 pagesBiology Investegatory Project 4saniya nanheNo ratings yet
- The Biology Project: Mendelian Genetics: Done By: B.Mithun Xii-ADocument28 pagesThe Biology Project: Mendelian Genetics: Done By: B.Mithun Xii-Apriya balaji88% (8)
- wdededDocument4 pageswdededsahooswapnaswarupNo ratings yet
- Gregor Johann Mendel: 20 July 1822-6 January 1884Document4 pagesGregor Johann Mendel: 20 July 1822-6 January 1884Yuki NakataNo ratings yet
- MedilianismDocument16 pagesMedilianismYujin AhnNo ratings yet
- Gregor Mendel and The Laws of HeredityDocument2 pagesGregor Mendel and The Laws of HeredityKawther Al mashaiNo ratings yet
- Mendelian GeneticsDocument49 pagesMendelian GeneticsKaiye ReitarNo ratings yet
- Biology Chapter 11Document26 pagesBiology Chapter 11zbettencourt409060No ratings yet
- RPP - Pewarisan Sifat Inggris Silabus RsbiDocument14 pagesRPP - Pewarisan Sifat Inggris Silabus RsbiAgus Joko SungkonoNo ratings yet
- Unit 6 Genetics and InheritanceDocument36 pagesUnit 6 Genetics and InheritancehanenalsukiniNo ratings yet
- Mendel and The Principles of HeredityDocument61 pagesMendel and The Principles of Heredityfae-ar_raziNo ratings yet
- Medelian GeneticsDocument4 pagesMedelian GeneticsKen PeñamanteNo ratings yet
- Technological Institute of The Philippines Quiapo, ManilaDocument4 pagesTechnological Institute of The Philippines Quiapo, ManilaKrystel LahomNo ratings yet
- Gregor Johann MendelDocument18 pagesGregor Johann Mendeldeepthi k thomas100% (1)
- Genetics BookDocument77 pagesGenetics BookelctoyasNo ratings yet
- Mam Sobia Slide 1Document15 pagesMam Sobia Slide 1www.shamiff.w32No ratings yet
- Lecture Note On Bio 411 (Population Genetics)Document8 pagesLecture Note On Bio 411 (Population Genetics)abuberkerabdullerhiiNo ratings yet
- The Beginnings of Genetics - REPORT BY STUDENTS OF 10ADocument18 pagesThe Beginnings of Genetics - REPORT BY STUDENTS OF 10AFawwazNo ratings yet
- Mendel Summarized ReviewerDocument4 pagesMendel Summarized ReviewerAstrid BacuñoNo ratings yet
- Mendel GregorDocument3 pagesMendel GregorTricia AvilaNo ratings yet
- Mendel_s_lowsDocument3 pagesMendel_s_lowsya708007No ratings yet
- Mendels Law of InheritanceDocument4 pagesMendels Law of InheritanceKriss ReissNo ratings yet
- A Research Paper On Gregor MendelDocument4 pagesA Research Paper On Gregor MendelUsairam SherazNo ratings yet
- Ka - Who Was Gregor MendelDocument6 pagesKa - Who Was Gregor Mendeljtymes713No ratings yet
- GENETICS-WPS OfficeDocument12 pagesGENETICS-WPS OfficeUTKARSH TAYALNo ratings yet
- Review Ee ErrDocument12 pagesReview Ee ErrMaryfleur BazarNo ratings yet
- BiologyDocument28 pagesBiology2w887q9kqjNo ratings yet
- CBSE Class - 12 - BiologyDocument7 pagesCBSE Class - 12 - BiologyAanav MittalNo ratings yet
- Mendelian GeneticsDocument23 pagesMendelian GeneticsPallavi Chaturvedi100% (1)
- Genetics 1st - 1Document90 pagesGenetics 1st - 1hdereje59No ratings yet
- Mendelian GeneticsDocument49 pagesMendelian GeneticsardhcyarNo ratings yet
- Principles of Inheritance and Variation - eMedicalPrepDocument12 pagesPrinciples of Inheritance and Variation - eMedicalPrepTUSHAR DASHNo ratings yet
- Mendel and His PeasDocument7 pagesMendel and His PeasMichael SopranoNo ratings yet
- Final Mendel Law 2Document9 pagesFinal Mendel Law 2ravip_29No ratings yet
- Module 4 CytogeneticsDocument21 pagesModule 4 CytogeneticsFrances Riane SimoyNo ratings yet
- Gregor Mendel The Father of GeneticsDocument2 pagesGregor Mendel The Father of GeneticsAnonymous BBs1xxk96VNo ratings yet
- Mendelian Concept of HeredityDocument15 pagesMendelian Concept of HereditySHEENA MAE DALGUNTASNo ratings yet
- Classical (Mendelian) GeneticsDocument32 pagesClassical (Mendelian) GeneticsKOMAL FATIMANo ratings yet
- Genetics June 18 2015Document71 pagesGenetics June 18 2015Didi AguiñagaNo ratings yet
- I Got It from My Mama! Gregor Mendel Explains Heredity - Science Book Age 9 | Children's Biology BooksFrom EverandI Got It from My Mama! Gregor Mendel Explains Heredity - Science Book Age 9 | Children's Biology BooksNo ratings yet
- VCXVCXXC FG Fhgfjdgfdedfxcfdfdffgfcvgdfgdghdcgvfdgbfhn GMJGHMKJHHDFDDFDGGFGGRFGDSGRFHGFJNHGKMJM, NNK MKJKHJK, M,.MMMM, KMKKKMKHGHM, NXZCDocument1 pageVCXVCXXC FG Fhgfjdgfdedfxcfdfdffgfcvgdfgdghdcgvfdgbfhn GMJGHMKJHHDFDDFDGGFGGRFGDSGRFHGFJNHGKMJM, NNK MKJKHJK, M,.MMMM, KMKKKMKHGHM, NXZCnfacmaNo ratings yet
- VCXVCXXC FG Fhgfjdgfdedfxcfdfdffgfcvgdfgdghdcgvfdgbfhn GMJGHMKJHHDFDDGGDSGRFHGFJNHGKMJM, NNKMKJKHJK, M,. MMMM, KMKKKMKHGHM, NDocument1 pageVCXVCXXC FG Fhgfjdgfdedfxcfdfdffgfcvgdfgdghdcgvfdgbfhn GMJGHMKJHHDFDDGGDSGRFHGFJNHGKMJM, NNKMKJKHJK, M,. MMMM, KMKKKMKHGHM, NnfacmaNo ratings yet
- VCXVCXXC FG Fhgfjdgfdedfgdfgdghdcgvfdgbfhngmjghmkjhhg DSGRFHGFJNHGKMJM, NNKMKJKHJK, M,.MMMM, KMKKKMKHGHM, NDocument1 pageVCXVCXXC FG Fhgfjdgfdedfgdfgdghdcgvfdgbfhngmjghmkjhhg DSGRFHGFJNHGKMJM, NNKMKJKHJK, M,.MMMM, KMKKKMKHGHM, NnfacmaNo ratings yet
- VCXVCXXC FG Fhgfjdgfdedfxcfdfdffgfcvgdfgdghdcgvfdgbfhn GMJGHMKJHHGDSGRFHGFJNHGKMJM, NNKMKJKHJK, M,.MMMM, K MKKKMKHGHM, NDocument1 pageVCXVCXXC FG Fhgfjdgfdedfxcfdfdffgfcvgdfgdghdcgvfdgbfhn GMJGHMKJHHGDSGRFHGFJNHGKMJM, NNKMKJKHJK, M,.MMMM, K MKKKMKHGHM, NnfacmaNo ratings yet
- VCXVCXXC FG Fhgfjdgfdedfxcfdfdffgfgdfgdghdcgvfdgbfhngm JGHMKJHHGDSGRFHGFJNHGKMJM, NNKMKJKHJK, M,.MMMM, KMK KKMKHGHM, NDocument1 pageVCXVCXXC FG Fhgfjdgfdedfxcfdfdffgfgdfgdghdcgvfdgbfhngm JGHMKJHHGDSGRFHGFJNHGKMJM, NNKMKJKHJK, M,.MMMM, KMK KKMKHGHM, NnfacmaNo ratings yet
- VCXVCXXC FG Fhgfjdgfdedfxcfdfdffgfcvgdfgdghdcgvfdgbfhn GMJGHMKJHHDFDDGGFGGRFGDSGRFHGFJNHGKMJM, NNKMK JKHJK, M,.MMMM, KMKKKMKHGHM, NXZCDocument1 pageVCXVCXXC FG Fhgfjdgfdedfxcfdfdffgfcvgdfgdghdcgvfdgbfhn GMJGHMKJHHDFDDGGFGGRFGDSGRFHGFJNHGKMJM, NNKMK JKHJK, M,.MMMM, KMKKKMKHGHM, NXZCnfacmaNo ratings yet
- VCXVCXXC FG Fhgfjdgfdedfxcfdfdffgfcvgdfgdghdcgvfdgbfhn GMJGHMKJHHDFDDFDGGFGGRFGDSGRFHGFJNHGKMJM, NNK MKJKVFVHJK, M,.MMMM, KMKKKMKHGHM, NXZCDocument1 pageVCXVCXXC FG Fhgfjdgfdedfxcfdfdffgfcvgdfgdghdcgvfdgbfhn GMJGHMKJHHDFDDFDGGFGGRFGDSGRFHGFJNHGKMJM, NNK MKJKVFVHJK, M,.MMMM, KMKKKMKHGHM, NXZCnfacmaNo ratings yet
- VCXVCXXC FG FHGFJDGFDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSG RFHGFJNHGKMJM, NNKMKJKHJK, M,.MMMM, KMKKKMKHGHM, NDocument1 pageVCXVCXXC FG FHGFJDGFDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSG RFHGFJNHGKMJM, NNKMKJKHJK, M,.MMMM, KMKKKMKHGHM, NnfacmaNo ratings yet
- VCXVCXXC FG Fhgfjdgfdedfxcfdgdfgdghdcgvfdgbfhngmjghmkj HHGDSGRFHGFJNHGKMJM, NNKMKJKHJK, M,.MMMM, KMKKKMKHGH M, NDocument1 pageVCXVCXXC FG Fhgfjdgfdedfxcfdgdfgdghdcgvfdgbfhngmjghmkj HHGDSGRFHGFJNHGKMJM, NNKMKJKHJK, M,.MMMM, KMKKKMKHGH M, NnfacmaNo ratings yet
- XC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJM, NNKMKJKHJK, M,.MMKMKKKMKHGHM, NDocument1 pageXC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJM, NNKMKJKHJK, M,.MMKMKKKMKHGHM, NnfacmaNo ratings yet
- VCXVCXXC FG Fhgfjdgfdedfxcfdfdffgfcvgdfgdghdcgvfdgbfhn GMJGHMKJHHDFDGDSGRFHGFJNHGKMJM, NNKMKJKHJK, M,.MM MM, KMKKKMKHGHM, NDocument1 pageVCXVCXXC FG Fhgfjdgfdedfxcfdfdffgfcvgdfgdghdcgvfdgbfhn GMJGHMKJHHDFDGDSGRFHGFJNHGKMJM, NNKMKJKHJK, M,.MM MM, KMKKKMKHGHM, NnfacmaNo ratings yet
- XC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJM, NNKMKJKHJK, M,.KMKKKMKHGHM, NDocument1 pageXC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJM, NNKMKJKHJK, M,.KMKKKMKHGHM, NnfacmaNo ratings yet
- XC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJKMKKKMKDocument1 pageXC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJKMKKKMKnfacmaNo ratings yet
- VCXVCXXC FG Fhgfjdgfdedfxcfdfdfgdfgdghdcgvfdgbfhngmjgh MKJHHGDSGRFHGFJNHGKMJM, NNKMKJKHJK, M,.MMMM, KMKKKM KHGHM, NDocument1 pageVCXVCXXC FG Fhgfjdgfdedfxcfdfdfgdfgdghdcgvfdgbfhngmjgh MKJHHGDSGRFHGFJNHGKMJM, NNKMKJKHJK, M,.MMMM, KMKKKM KHGHM, NnfacmaNo ratings yet
- XC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJM, NNKMKJKHJK, M,.MMMM, KMKKKMKHGHM, NDocument1 pageXC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJM, NNKMKJKHJK, M,.MMMM, KMKKKMKHGHM, NnfacmaNo ratings yet
- XC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJM, NNKMKKKMKHGHM, NDocument1 pageXC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJM, NNKMKKKMKHGHM, NnfacmaNo ratings yet
- XC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJM, NNKMKJKHJK, MKMKKKMKHGHM, NDocument1 pageXC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJM, NNKMKJKHJK, MKMKKKMKHGHM, NnfacmaNo ratings yet
- DFBFGBFV 10 CVDFVCDFVFDVFDVFDFRGRHRJMHJK, KJ, JK, KJM, KJHKJKFDVDGVVVVVVVVVVVVVVVVVVVVVVVVVVVVV Vvfddscefdgru, JMNHBVCX, Kmujnybhtgvrfdli, KmujnhbgDocument1 pageDFBFGBFV 10 CVDFVCDFVFDVFDVFDFRGRHRJMHJK, KJ, JK, KJM, KJHKJKFDVDGVVVVVVVVVVVVVVVVVVVVVVVVVVVVV Vvfddscefdgru, JMNHBVCX, Kmujnybhtgvrfdli, KmujnhbgnfacmaNo ratings yet
- XC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJKMKKKMKHGHDocument1 pageXC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJKMKKKMKHGHnfacmaNo ratings yet
- XC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJKMKKKMKHGHM, NDocument1 pageXC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJKMKKKMKHGHM, NnfacmaNo ratings yet
- XC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJM, NNKMKJKHJKMKKKMKHGHM, NDocument1 pageXC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJM, NNKMKJKHJKMKKKMKHGHM, NnfacmaNo ratings yet
- XC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJKMKKDocument1 pageXC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJKMKKnfacmaNo ratings yet
- XC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJM, NNKMKKMKKKMKHGHM, NDocument1 pageXC FGFHGFJDG FDGDFGDGHDCGVFDGBFHNGMJGHMKJHHGDSGRFHGFJN HGKMJM, NNKMKKMKKKMKHGHM, NnfacmaNo ratings yet
- DFBFGBFV 10 CVDFVCDFVFDVFDVFDFRGRHRJMHJK, KJ, JK, KJM, KJHKJKFDVDGVVVVVVVVVVVVVVVVVVVVVVVVVVVVV Vvfddscefdgrdfdsfdsfdsu, JMNHBVCX, Kmujnybhtgvrfdli, KmujnhbgDocument1 pageDFBFGBFV 10 CVDFVCDFVFDVFDVFDFRGRHRJMHJK, KJ, JK, KJM, KJHKJKFDVDGVVVVVVVVVVVVVVVVVVVVVVVVVVVVV Vvfddscefdgrdfdsfdsfdsu, JMNHBVCX, Kmujnybhtgvrfdli, KmujnhbgnfacmaNo ratings yet
- DFBFGBFV 10 CVDFVCDFVFDVFDVFDFRGRHRJMHJK, KJ, JK, KJM, KJHKJKFDVDGVVVVVVVVVVVVVVVVVVVVVVVVVVVVV Vvfddscefdgrdfdsfdsfdsu, JMNHBVCX, Kmujnybhtgvrfdli, Kmuwedwfdfdjnhbgdsfsfsfcdscf DscfsddfscdscdsvsDocument1 pageDFBFGBFV 10 CVDFVCDFVFDVFDVFDFRGRHRJMHJK, KJ, JK, KJM, KJHKJKFDVDGVVVVVVVVVVVVVVVVVVVVVVVVVVVVV Vvfddscefdgrdfdsfdsfdsu, JMNHBVCX, Kmujnybhtgvrfdli, Kmuwedwfdfdjnhbgdsfsfsfcdscf DscfsddfscdscdsvsnfacmaNo ratings yet
- DFBFGBFV 10 CVDFVCDFVFDVFDVFDFRGRHRJMHJK, KJ, JK, KJM, KJHKJKFDVDGVVVVVVVVVVVVVVVVVVVVVVVVVVVVV Vvfdu, JMNHBVCX, Kmujnybhtgvrfdli, KmujnhbgDocument1 pageDFBFGBFV 10 CVDFVCDFVFDVFDVFDFRGRHRJMHJK, KJ, JK, KJM, KJHKJKFDVDGVVVVVVVVVVVVVVVVVVVVVVVVVVVVV Vvfdu, JMNHBVCX, Kmujnybhtgvrfdli, KmujnhbgnfacmaNo ratings yet
- DFBFGBFV 10 CVDFVCDFVFDVFDVFDFRGRHRJMHJK, KJ, JK, KJM, Kjhkjku, JMNHBVCX, Kmujnybhtgvrfdli, KmujnhbgDocument1 pageDFBFGBFV 10 CVDFVCDFVFDVFDVFDFRGRHRJMHJK, KJ, JK, KJM, Kjhkjku, JMNHBVCX, Kmujnybhtgvrfdli, KmujnhbgnfacmaNo ratings yet
- DFBFGBFV 10Document1 pageDFBFGBFV 10nfacmaNo ratings yet
- DFBFGBFV 10 CVDFVCDFVFDVFDVFDFRGRHRJMHJK, KJ, JK, KJM, KJHKJKFDVDGVVVVVVVVVVVVVVVVVVVVVVVVVVVVV Vvfddscefdgrdfdsfdsfdsu, JMNHBVCX, Kmujnybhtgvrfdli, Kmuwedwfdfdjnhbgdsfsfsfcdscf DSCFSDDocument1 pageDFBFGBFV 10 CVDFVCDFVFDVFDVFDFRGRHRJMHJK, KJ, JK, KJM, KJHKJKFDVDGVVVVVVVVVVVVVVVVVVVVVVVVVVVVV Vvfddscefdgrdfdsfdsfdsu, JMNHBVCX, Kmujnybhtgvrfdli, Kmuwedwfdfdjnhbgdsfsfsfcdscf DSCFSDnfacmaNo ratings yet
- Nutrition in Aging: Nurpudji A. Taslim Nutrition Department School of Medicine Hasanuddin University at 2005Document27 pagesNutrition in Aging: Nurpudji A. Taslim Nutrition Department School of Medicine Hasanuddin University at 2005nfacmaNo ratings yet
- Monster Genetics LabDocument4 pagesMonster Genetics LabJoan LuisNo ratings yet
- Learning Activity Report 2Document18 pagesLearning Activity Report 2Mikee GarzonNo ratings yet
- Drosophila MelanogasterDocument6 pagesDrosophila Melanogasterdanielle_wolbert100% (1)
- Evolution Inquiry Peppered MothsDocument8 pagesEvolution Inquiry Peppered Mothsapi-394564312No ratings yet
- Genetics AssignmentDocument20 pagesGenetics AssignmentPérvéz NázírNo ratings yet
- Jose P. Laurel Sr. High SchoolDocument8 pagesJose P. Laurel Sr. High SchoolEricha SolomonNo ratings yet
- Evolutionary Limits and Constraints: Ary HoffmannDocument6 pagesEvolutionary Limits and Constraints: Ary HoffmannIsaac BismonteNo ratings yet
- Study Materials: Vedantu Innovations Pvt. Ltd. Score High With A Personal Teacher, Learn LIVE Online!Document8 pagesStudy Materials: Vedantu Innovations Pvt. Ltd. Score High With A Personal Teacher, Learn LIVE Online!sushantNo ratings yet
- Heterogeneity in Statistical Genetics: Derek Gordon Stephen J. Finch Wonkuk KimDocument366 pagesHeterogeneity in Statistical Genetics: Derek Gordon Stephen J. Finch Wonkuk KimWulil AlbabNo ratings yet
- Use of Linear Algebra in Autosomal Inheritance/ BioinformaticsDocument4 pagesUse of Linear Algebra in Autosomal Inheritance/ Bioinformaticsshiksha torooNo ratings yet
- 6 Genetics PDFDocument97 pages6 Genetics PDFasaNo ratings yet
- Bio 101 QuestionDocument15 pagesBio 101 QuestionMohammed kashimNo ratings yet
- UCLA Genetics LS4 - Midterm 1 Spr14 KeyDocument9 pagesUCLA Genetics LS4 - Midterm 1 Spr14 KeyMari JaneNo ratings yet
- SCI - Learning Packet - Biology Week 3Document9 pagesSCI - Learning Packet - Biology Week 3Pal, Julia HanneNo ratings yet
- Genetics Study Guide AnswersDocument23 pagesGenetics Study Guide Answersapi-366982317No ratings yet
- Assignment 4 ORIDocument2 pagesAssignment 4 ORIK WATHSALANo ratings yet
- Multiple Choice: Choose Best (2.5 Points Each, 80Document7 pagesMultiple Choice: Choose Best (2.5 Points Each, 80Florence BamigbolaNo ratings yet
- BioPsych Exam 1 Study GuideDocument5 pagesBioPsych Exam 1 Study GuideShiv PatelNo ratings yet
- Biology Form 4 NotesDocument52 pagesBiology Form 4 Notesmohamed ibrahim osman100% (4)
- Mariah Q. - Revised Genetic Mystery Pixel Art - GeneticsDocument2 pagesMariah Q. - Revised Genetic Mystery Pixel Art - GeneticsMariah QuayesonNo ratings yet
- Biology a U8 Sample WorkDocument31 pagesBiology a U8 Sample Workcarternoble408No ratings yet
- Jibachha Animal Husbandry PDFDocument17 pagesJibachha Animal Husbandry PDFJibachha ShahNo ratings yet
- Psychological GeneticsDocument24 pagesPsychological GeneticsdevNo ratings yet
- Project Heredity & DNA v3Document13 pagesProject Heredity & DNA v3maxvandorsser07No ratings yet
- Pre TestDocument2 pagesPre TestJane GohNo ratings yet
- Punnett Square: Ms. Elaine B. EspejonDocument24 pagesPunnett Square: Ms. Elaine B. EspejonElaineNo ratings yet
- Dragon Genetics ProtocolDocument5 pagesDragon Genetics ProtocolDanielle MedinaNo ratings yet
- Heredity (Class 10) NotesDocument4 pagesHeredity (Class 10) NotesAlok Naik100% (1)