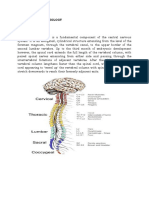
Anatomia Si Localizare in Boli Spinale
Anatomia Si Localizare in Boli Spinale
Download as doc, pdf, or txt
You might also like
- C21 UM enUS V2.2Document76 pagesC21 UM enUS V2.2Fernando de pedroNo ratings yet
- (Neurophysiology) (EEG) Atlas of EEG and Seizure SemiologyDocument211 pages(Neurophysiology) (EEG) Atlas of EEG and Seizure SemiologyHalit Diana83% (6)
- Diagnosis Imaging For The Emergency PhysicianDocument911 pagesDiagnosis Imaging For The Emergency PhysicianHalit Diana90% (10)
- Spinal Cord: StructureDocument5 pagesSpinal Cord: StructureIka LestariNo ratings yet
- Spinal Cord Quick SummaryDocument5 pagesSpinal Cord Quick Summarykaykay200058No ratings yet
- Spinal Cord: Navigation SearchDocument16 pagesSpinal Cord: Navigation SearchritzchoiNo ratings yet
- Biology ReportDocument3 pagesBiology ReportzyrianNo ratings yet
- Spinal Cord LesionsDocument110 pagesSpinal Cord Lesionsgtaha80100% (2)
- Saraf Spinal: Neuroanatomy, Somatic Nervous SystemDocument10 pagesSaraf Spinal: Neuroanatomy, Somatic Nervous SystemFitri SukmawatiNo ratings yet
- Somatosensory PathwaysDocument12 pagesSomatosensory PathwaysDimasBanyakRejekiNo ratings yet
- Spinal CordDocument12 pagesSpinal Cordharis13.hsNo ratings yet
- Spinal Cord: Achmad AminuddinDocument29 pagesSpinal Cord: Achmad AminuddinSurajul AudaNo ratings yet
- Anatomy of Spinal CordDocument33 pagesAnatomy of Spinal CordAmy BabynotNo ratings yet
- SCI Management ExtrasDocument2 pagesSCI Management ExtrasLoredana TaclitNo ratings yet
- Lab 17 The Spinal Cord Somatic NervesDocument18 pagesLab 17 The Spinal Cord Somatic Nervesrhycelayon304No ratings yet
- Anatomy SpineDocument6 pagesAnatomy SpineKris AdiNo ratings yet
- Central and Peripheral Nervous System, Spinal Cord and Spinal NervesDocument4 pagesCentral and Peripheral Nervous System, Spinal Cord and Spinal NervesDaniel DanielNo ratings yet
- Neuroanatomy 1Document65 pagesNeuroanatomy 1maNo ratings yet
- Spinal Cord and Spinal Nerves ReviewerDocument5 pagesSpinal Cord and Spinal Nerves ReviewerTay LenaNo ratings yet
- Spinal Cord, Ascending PathwayDocument52 pagesSpinal Cord, Ascending PathwayKolawole Adesina KehindeNo ratings yet
- Cabatbat, Sable A. BSN Iv - D: Brain MusclesDocument4 pagesCabatbat, Sable A. BSN Iv - D: Brain MusclesOdyNo ratings yet
- Iv. The Spinal Cord: Cordotomy Lumbar PunctureDocument0 pagesIv. The Spinal Cord: Cordotomy Lumbar PunctureAnonymous t5TDwdNo ratings yet
- Chapter #5: The Spinal CordDocument4 pagesChapter #5: The Spinal CordRazvanNo ratings yet
- Autonomic Nervous System: Aqsa Khan (MPT Sem)Document19 pagesAutonomic Nervous System: Aqsa Khan (MPT Sem)furquan 810No ratings yet
- ue 1-Broc s Are ʹ an area of the frontal lobe, usually in the left hemisphere, that directs the muscle movements involvedDocument31 pagesue 1-Broc s Are ʹ an area of the frontal lobe, usually in the left hemisphere, that directs the muscle movements involvedMhmd IrakyNo ratings yet
- The Spinal Cord: Lec-11-Dr - Maysoon AlkazzazDocument12 pagesThe Spinal Cord: Lec-11-Dr - Maysoon AlkazzazRose ParisNo ratings yet
- ЦНС - 2019Document50 pagesЦНС - 2019bekafop813No ratings yet
- Spine Structure and FunctionDocument19 pagesSpine Structure and Functionakhil dattaNo ratings yet
- Spinal Cord AnatomyDocument34 pagesSpinal Cord AnatomyYana CatherineNo ratings yet
- Anatomy IV Lab ManualDocument44 pagesAnatomy IV Lab ManualTanish KumarNo ratings yet
- Spinal Nerves & PluxesDocument31 pagesSpinal Nerves & PluxesJyoti SidhuNo ratings yet
- 18 - Cervical SympatheticDocument10 pages18 - Cervical SympatheticEyma SuldamadaNo ratings yet
- Anatomy and Physiology The Spinal CordDocument11 pagesAnatomy and Physiology The Spinal CordKatrina CaveNo ratings yet
- What Are Spinal Tracts?Document2 pagesWhat Are Spinal Tracts?Ahsan JamilNo ratings yet
- Control Body Movements and FunctionsDocument7 pagesControl Body Movements and Functionsnnce eceNo ratings yet
- Overview of Spinal NervesDocument12 pagesOverview of Spinal NervesGrace VersoniNo ratings yet
- Somatosensory PathwaysDocument36 pagesSomatosensory PathwaysAnonymous w3vYureNo ratings yet
- Spinal CordDocument35 pagesSpinal CordZeenat Arshad 6624No ratings yet
- Katya Anaphysio NewDocument6 pagesKatya Anaphysio NewKriska Herrero TumamakNo ratings yet
- Lab. II. Spinal CordDocument7 pagesLab. II. Spinal CordAun CanineNo ratings yet
- Asynch NotesDocument18 pagesAsynch NotesEricka Mae BruanNo ratings yet
- Autonomic N.SDocument7 pagesAutonomic N.SAjigotto ubiq.No ratings yet
- Npte - Neuroanatomy Lec 2Document31 pagesNpte - Neuroanatomy Lec 2Densen Raphael C. Antonio100% (2)
- 10 - Spinal Cord Injuries Lec.1Document7 pages10 - Spinal Cord Injuries Lec.1AmmarNo ratings yet
- Spinal CordDocument27 pagesSpinal Cordsiddharth.roy26587No ratings yet
- Nervous PathwaysDocument20 pagesNervous Pathwaysdod_nurNo ratings yet
- تشريح عام 4Document22 pagesتشريح عام 4tabarkarkan596No ratings yet
- Anatomy and Physiology of The Spine: Google ImagesDocument9 pagesAnatomy and Physiology of The Spine: Google Imagesraquel maniegoNo ratings yet
- Spinal Cord and MedullaDocument34 pagesSpinal Cord and MedullaveegeerNo ratings yet
- Spinal CordDocument38 pagesSpinal CordAsad OsmanNo ratings yet
- The Nervous SystemDocument40 pagesThe Nervous Systememmy9ize1No ratings yet
- Spinal NervesDocument15 pagesSpinal NervesPrachNo ratings yet
- MODULE 3 SGD Autonomic Nervous SystemDocument6 pagesMODULE 3 SGD Autonomic Nervous SystemKristine Jade OdtujanNo ratings yet
- 1 - Spinal Cord NotesDocument7 pages1 - Spinal Cord NotesOceanChan QdNo ratings yet
- #Anatomy of The Spinal CordDocument15 pages#Anatomy of The Spinal CordMaryamNo ratings yet
- Spinal ChordDocument8 pagesSpinal ChordmalovealbeniaNo ratings yet
- Continuum Spinal Cord AnatomyDocument25 pagesContinuum Spinal Cord AnatomyslojnotakNo ratings yet
- Spinal NerveDocument7 pagesSpinal NerveGhazi Uddin AhmedNo ratings yet
- 12 Spinal Cord TractsDocument39 pages12 Spinal Cord Tractssjs6r8wwv9No ratings yet
- Summary of Stanley Rosenberg's Accessing the Healing Power of the Vagus NerveFrom EverandSummary of Stanley Rosenberg's Accessing the Healing Power of the Vagus NerveNo ratings yet
- Neuromodulation in Headache and Facial Pain Management: Principles, Rationale and Clinical DataFrom EverandNeuromodulation in Headache and Facial Pain Management: Principles, Rationale and Clinical DataGiorgio LambruNo ratings yet
- Anatomia Si Localizare in Boli SpinaleDocument8 pagesAnatomia Si Localizare in Boli SpinaleHalit DianaNo ratings yet
- Boli Vertebro MedulareDocument12 pagesBoli Vertebro MedulareHalit DianaNo ratings yet
- Solution Manual For Basic Robotics 1st Edition by Dinwiddie ISBN 9781133950196 9781133950196Document10 pagesSolution Manual For Basic Robotics 1st Edition by Dinwiddie ISBN 9781133950196 9781133950196johnNo ratings yet
- ReferatDocument7 pagesReferatMuh Arya SaputraNo ratings yet
- 5 - ArthrologyDocument26 pages5 - Arthrologydrshubhamvyas.08No ratings yet
- The ChosenDocument2 pagesThe ChosenBigFriendlyKittyCatNo ratings yet
- MRCS Osce Sample-2 PDFDocument25 pagesMRCS Osce Sample-2 PDFAmmarNo ratings yet
- Accreditation No.: 1030-091918-093Document4 pagesAccreditation No.: 1030-091918-093Leo Buquiran AcabalNo ratings yet
- Part of SpeechDocument77 pagesPart of SpeechGarnis NurfadilaNo ratings yet
- Owner'S Manual: Automatic Positive Locking Traction DifferentialDocument24 pagesOwner'S Manual: Automatic Positive Locking Traction Differentialhelp3r0% (1)
- A Report On The 35 Border Collies From Flat Creek in Sprakers, NYDocument36 pagesA Report On The 35 Border Collies From Flat Creek in Sprakers, NYCaitlin Mason100% (1)
- Hernia Ebook FullDocument24 pagesHernia Ebook FullMiguelon GarciaNo ratings yet
- SUN2000 - (8KTL-20KTL) - M2 User ManualDocument136 pagesSUN2000 - (8KTL-20KTL) - M2 User ManualMuresan AlinaNo ratings yet
- Your Guide To Sun SalutationsDocument3 pagesYour Guide To Sun SalutationsIswarya Ravi100% (1)
- Avulsed TeethDocument3 pagesAvulsed TeethSimona DobreNo ratings yet
- Mandibular FractresDocument37 pagesMandibular Fractresdileep900No ratings yet
- Family Sues Administration After Daughter Gets Beat Up at SchoolDocument16 pagesFamily Sues Administration After Daughter Gets Beat Up at SchoolJeff WyattNo ratings yet
- Full Sectional Anatomy by MRI and CT 3rd Edition Georges Y. El-Khoury Ebook All ChaptersDocument84 pagesFull Sectional Anatomy by MRI and CT 3rd Edition Georges Y. El-Khoury Ebook All Chaptersmupeukhatz100% (3)
- Inglessis DeclarationDocument3 pagesInglessis DeclarationTHROnlineNo ratings yet
- New Qs W AnaswerDocument50 pagesNew Qs W AnaswerHengameh Javahery100% (1)
- Sports Safety: Related Kidshealth LinksDocument6 pagesSports Safety: Related Kidshealth LinksIrine ReyesNo ratings yet
- Principles in Trauma ManagementDocument34 pagesPrinciples in Trauma Managementvirz23No ratings yet
- Fall Protection Rescue Plan ProcedureDocument3 pagesFall Protection Rescue Plan ProcedureDean Delon NanlohyNo ratings yet
- Anatomical Position of The BodyDocument5 pagesAnatomical Position of The BodyPrecious MapehNo ratings yet
- Final Syllabus: Anatomy Upper and Lower LimbDocument6 pagesFinal Syllabus: Anatomy Upper and Lower LimbSbahat FatimaNo ratings yet
- Lower Leg and FootDocument40 pagesLower Leg and Footapi-295870055No ratings yet
- Rebar - WikipediaDocument12 pagesRebar - WikipediaglennNo ratings yet
- Shoulder Rehab Ver2Document7 pagesShoulder Rehab Ver2Daniel Hernandez Tellerias100% (2)
- Accessory Nerve - Cranial Nerve XI AtfDocument3 pagesAccessory Nerve - Cranial Nerve XI AtfCathy SolivarNo ratings yet
- Anterior Knee Pain ExercisesDocument6 pagesAnterior Knee Pain ExercisesJobin VargheseNo ratings yet
- Guide To Speaking and ListeningDocument1 pageGuide To Speaking and ListeningSrna GurgurNo ratings yet