Alpaca With Gastrointestinal Parasites
Alpaca With Gastrointestinal Parasites
Download as pdf or txt
You might also like
- MycoViro ExamDocument6 pagesMycoViro Examjaypster30No ratings yet
- ch01 Lecture PPT A1-1Document43 pagesch01 Lecture PPT A1-1Leona Rabe100% (10)
- Trabajo MichiganDocument10 pagesTrabajo MichiganJuan Martín VargasNo ratings yet
- Small Rum. 21Document8 pagesSmall Rum. 21hrithikecNo ratings yet
- Coles ResistanceDocument3 pagesColes Resistancehorpor17No ratings yet
- O-073 Seroprevalence of Enzootic Abortion and Toxoplasmosis in Sheep Flocks Experiencing Reproductive LossesDocument2 pagesO-073 Seroprevalence of Enzootic Abortion and Toxoplasmosis in Sheep Flocks Experiencing Reproductive LossesAbidas ArbNo ratings yet
- Case Study 2Document9 pagesCase Study 2Rowena G. TenecioNo ratings yet
- Philipp J Vet Anim Sci 2013, 39 (1) : 99-106Document8 pagesPhilipp J Vet Anim Sci 2013, 39 (1) : 99-106Rezki WidiansyahNo ratings yet
- Five Years of Partula Snail Pre-Export Health ScreeningDocument12 pagesFive Years of Partula Snail Pre-Export Health ScreeningJaviera Meza LunaNo ratings yet
- AnaplasmaDocument10 pagesAnaplasmabryanpes71No ratings yet
- Fda ST398Document30 pagesFda ST398ophexus1205No ratings yet
- Declaran Dias No Laborables en El Sector Publico Correspondi Decreto Supremo N 002 2019 PCM 1728509 1Document6 pagesDeclaran Dias No Laborables en El Sector Publico Correspondi Decreto Supremo N 002 2019 PCM 1728509 1JACKYNICENo ratings yet
- Phage Therapy: The Escherichia Coli Experience: ReviewDocument8 pagesPhage Therapy: The Escherichia Coli Experience: ReviewAbhay Kumar MohiniNo ratings yet
- Evaluación de Campo de Dos Fasciolicidas en Bovinos Rubia Gallega.Document62 pagesEvaluación de Campo de Dos Fasciolicidas en Bovinos Rubia Gallega.Raul alejandro Kim gomezNo ratings yet
- Giardia Duodenalis Trophozoites Isolated From ADocument16 pagesGiardia Duodenalis Trophozoites Isolated From AluismitlvNo ratings yet
- Kelinci PDFDocument11 pagesKelinci PDFmedeNo ratings yet
- Journal of Veterinary Medicine Series A - 2003 - Herradora - Effect of Oral Enrofloxacin and Florfenicol on Pigs (1)Document5 pagesJournal of Veterinary Medicine Series A - 2003 - Herradora - Effect of Oral Enrofloxacin and Florfenicol on Pigs (1)emmanouelaapostolopoulouNo ratings yet
- Peer Reviewed: An Irish Perspective On CryptosporidiumDocument6 pagesPeer Reviewed: An Irish Perspective On CryptosporidiumDrivailaNo ratings yet
- Enzootic Abortion in Ewes: A Review of Recent Developments in DiagnosticsDocument3 pagesEnzootic Abortion in Ewes: A Review of Recent Developments in DiagnosticsErnesto VersailleNo ratings yet
- CA HA LA MRSA Frontiers in MicrobiologyDocument7 pagesCA HA LA MRSA Frontiers in MicrobiologyAmanda SilvaNo ratings yet
- Colistin Resistance in Escherichia Coli and Klebsiella EcuadorDocument8 pagesColistin Resistance in Escherichia Coli and Klebsiella EcuadorALVARO ESNEIDER AVILA GUZMANNo ratings yet
- The Efficacy of Vapco Cypermethrin Against Cattle Ticks in TanzaniaDocument8 pagesThe Efficacy of Vapco Cypermethrin Against Cattle Ticks in TanzaniapetnagagiNo ratings yet
- Self-Cure of Trypanosoma Brucei Brucei Infection in West African Dwarf SheepDocument7 pagesSelf-Cure of Trypanosoma Brucei Brucei Infection in West African Dwarf SheepBishop David SandaNo ratings yet
- Antiparasitic Activity of An Ivermectin and PraziqDocument16 pagesAntiparasitic Activity of An Ivermectin and PraziqPablo OrricoNo ratings yet
- 26 - Gastrophilus PecorumDocument3 pages26 - Gastrophilus PecorumHafiz Zamad Ur RehmanNo ratings yet
- Anthelmintic Resistance in Cattle Nematodes in The USDocument9 pagesAnthelmintic Resistance in Cattle Nematodes in The USNany ArizaNo ratings yet
- REVIEWA Naplasma Marginale and Anaplasma Phagocytophilum-Rickettsiales Pathogens of Veterinary and Publichealth SignificanceDocument17 pagesREVIEWA Naplasma Marginale and Anaplasma Phagocytophilum-Rickettsiales Pathogens of Veterinary and Publichealth Significanceyudhi arjentiniaNo ratings yet
- Influence of Subtherapeutic Levels of Oxytetacycline On Salmonella Typhimurim in Swine, Calves, and ChickensDocument9 pagesInfluence of Subtherapeutic Levels of Oxytetacycline On Salmonella Typhimurim in Swine, Calves, and ChickensLa Fille D'ÈveNo ratings yet
- Selective DefecationDocument12 pagesSelective Defecation1anna.waffelNo ratings yet
- Herds of Udayapur 2063: in MuleDocument34 pagesHerds of Udayapur 2063: in MuleDr.Kedar Karki ,M.V.Sc.Preventive Vet.Medicine CLSU PhilippinesNo ratings yet
- Veterinary Parasitology: Research PaperDocument5 pagesVeterinary Parasitology: Research PaperDr-Sadaqat Ali RaoNo ratings yet
- Giardia Duodenalis and Cryptosporidium Spp. in A Veterinary College Bovine Teaching HerdDocument7 pagesGiardia Duodenalis and Cryptosporidium Spp. in A Veterinary College Bovine Teaching HerdluismitlvNo ratings yet
- Parasite Control in Pasture-Grazed Dairy Cattle: Are We at The Edge of A Precipice?Document7 pagesParasite Control in Pasture-Grazed Dairy Cattle: Are We at The Edge of A Precipice?Oktrestu Dwi PutraNo ratings yet
- Gastrointestinal Protozoa in Non-human Primates of Four Zoological Garden in BelgiumDocument11 pagesGastrointestinal Protozoa in Non-human Primates of Four Zoological Garden in BelgiummateusNo ratings yet
- Lezama-Gutiérrez Et Al. - 2005 - Susceptibility of Biological Stages of The Horn FLDocument8 pagesLezama-Gutiérrez Et Al. - 2005 - Susceptibility of Biological Stages of The Horn FLJuan UrrutiaNo ratings yet
- Duration of Protective Immunity Against Ovine Haemonchosis Following Vaccination With The Nematode Gut Membrane Antigen H L LDocument5 pagesDuration of Protective Immunity Against Ovine Haemonchosis Following Vaccination With The Nematode Gut Membrane Antigen H L LAngéllica MaríaNo ratings yet
- Food Control: Lai Kuan Tan, Peck Toung Ooi, Kwai Lin ThongDocument7 pagesFood Control: Lai Kuan Tan, Peck Toung Ooi, Kwai Lin ThongMafer JacomeNo ratings yet
- Interactive Effects of Turkeypox Virus and Plasmodium Hermani On Turkey PoultsDocument8 pagesInteractive Effects of Turkeypox Virus and Plasmodium Hermani On Turkey PoultsHeryansyah SkhNo ratings yet
- Art 07Document8 pagesArt 07Alejandro Acuña MellaNo ratings yet
- Juvenile Mortality in Captive Lesser Kudu (Tragelaphus Imberbis) at Basle Zoo and Is Relation To Nutrition and HusbandryDocument7 pagesJuvenile Mortality in Captive Lesser Kudu (Tragelaphus Imberbis) at Basle Zoo and Is Relation To Nutrition and Husbandrydra.ozonioNo ratings yet
- PublishedDocument5 pagesPublishedErnesto VersailleNo ratings yet
- 1-s2.0-S0304401707006632-mainDocument10 pages1-s2.0-S0304401707006632-mainmfuentesNo ratings yet
- Lima Barbero2019Document10 pagesLima Barbero2019spin_echoNo ratings yet
- (Paper) Prevalence of an inflammation & necrosis syndrome in suckling piglets - 2019年Document11 pages(Paper) Prevalence of an inflammation & necrosis syndrome in suckling piglets - 2019年Matthew HsuNo ratings yet
- Factors Affecting Fecal Egg Counts in Periparturient Katahdin Ewes and Their LambDocument10 pagesFactors Affecting Fecal Egg Counts in Periparturient Katahdin Ewes and Their LambJesus ekNo ratings yet
- Desease Control On GoatsDocument14 pagesDesease Control On Goats180006 RosmilahNo ratings yet
- Wormcontrol AHR Smallholder Sheep FarmsDocument7 pagesWormcontrol AHR Smallholder Sheep FarmsAfework KetemaNo ratings yet
- Vences A134 PDFDocument12 pagesVences A134 PDFLucìa SolerNo ratings yet
- AspergillusDocument15 pagesAspergillusAbdullateef AbdulmusawwirNo ratings yet
- ASPERGILLUSDocument27 pagesASPERGILLUSAbdullateef AbdulmusawwirNo ratings yet
- Ent Rise Ad C: S. Andrews, T N. J. K. Hole,? E. A. Munn$ Andi T. P. RolphifiDocument8 pagesEnt Rise Ad C: S. Andrews, T N. J. K. Hole,? E. A. Munn$ Andi T. P. RolphifiAngéllica MaríaNo ratings yet
- 01-PEA-JVS-50-2Document13 pages01-PEA-JVS-50-2lhernandez311No ratings yet
- Vancomycin Resistant EnterococciDocument238 pagesVancomycin Resistant EnterococciNoha Alaa EldinNo ratings yet
- The EpidemologyDocument15 pagesThe EpidemologyLakshmiNarayana Reddy BynagariNo ratings yet
- Rathinasamy 2018Document8 pagesRathinasamy 2018Krešimir SeverinNo ratings yet
- stear2001Document7 pagesstear2001Esteban CastañoNo ratings yet
- Prevalence of Gastrointestinal ParasitesDocument6 pagesPrevalence of Gastrointestinal ParasitesdharnfestusbNo ratings yet
- AMR Profiles of Common Mastitis Pathogens On Canadian Dairy FarmsDocument14 pagesAMR Profiles of Common Mastitis Pathogens On Canadian Dairy FarmsApril LarasatiNo ratings yet
- Oecophorine Genera of Australia I: The Wingia Group (Lepidoptera: Oecophoridae)From EverandOecophorine Genera of Australia I: The Wingia Group (Lepidoptera: Oecophoridae)No ratings yet
- Full Blast PLUS 1 (Beginners) - Grammar Test - Mod 2Document2 pagesFull Blast PLUS 1 (Beginners) - Grammar Test - Mod 2Atlanticcca0% (1)
- Sea LifeDocument8 pagesSea Lifelaureblct23No ratings yet
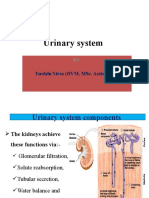
- Urinary System: Tsedalu Yirsa (DVM, MSC, Assist Prof.)Document18 pagesUrinary System: Tsedalu Yirsa (DVM, MSC, Assist Prof.)Mohammed OumerNo ratings yet
- PDF of Daddy Is Mine Veranna Leonand Full Chapter EbookDocument69 pagesPDF of Daddy Is Mine Veranna Leonand Full Chapter Ebookpearlfulae90100% (11)
- Biochemistry of Groundnut Synthesis and Its Chemical CompositionDocument23 pagesBiochemistry of Groundnut Synthesis and Its Chemical CompositionsushmitaNo ratings yet
- Core Competencies-Perform Pre Lay and Lay Activities CO Ms. DianDocument6 pagesCore Competencies-Perform Pre Lay and Lay Activities CO Ms. Dianrobelyn veranoNo ratings yet
- Lupin Seed As Replacement For Fishmeal in African Catfish (Clarias Gariepienus) FeedDocument26 pagesLupin Seed As Replacement For Fishmeal in African Catfish (Clarias Gariepienus) FeedBashir Muzakkar100% (1)
- PDF ViewDocument5 pagesPDF ViewAbhishek Singh SikarwarNo ratings yet
- Checking Harvesting Tools, Net Fish Cages and FramesDocument24 pagesChecking Harvesting Tools, Net Fish Cages and FramesClark Vergara CabaralNo ratings yet
- Efficacy of Physical Therapy On Cervical Muscle ActivityDocument9 pagesEfficacy of Physical Therapy On Cervical Muscle Activityrudhras22No ratings yet
- BrochureDocument5 pagesBrochureVoltaire Delos ReyesNo ratings yet
- Use Manual: 24k Golden Beauty Bar-Fashion Ideas Originate From JapanDocument2 pagesUse Manual: 24k Golden Beauty Bar-Fashion Ideas Originate From JapanJayvee MirandaNo ratings yet
- Blood Report TateDocument4 pagesBlood Report TateGina Watkins100% (2)
- Dichotomous Key For HistologyDocument3 pagesDichotomous Key For HistologyJeon JungkookieNo ratings yet
- Neet Structural Organisation in Animals TestDocument3 pagesNeet Structural Organisation in Animals TestShrishNo ratings yet
- Vet Tech Research Paper TopicsDocument8 pagesVet Tech Research Paper Topicsgepuz0gohew2100% (1)
- Third Eye SpellDocument2 pagesThird Eye SpellRudrani Sarkar100% (2)
- Hematology: AbbreviationsDocument5 pagesHematology: AbbreviationsReyven Niña DyNo ratings yet
- HJMXN Handbook - Of.evolution..the - Evolution.of - Living.systemsDocument240 pagesHJMXN Handbook - Of.evolution..the - Evolution.of - Living.systemsОюундэлгэрэй Батчулуун100% (5)
- Muntu, The Intelligent OneDocument5 pagesMuntu, The Intelligent OneDeryl ArtsNo ratings yet
- Soal Bahasa Inggris Paket C Setara SMA/MADocument22 pagesSoal Bahasa Inggris Paket C Setara SMA/MApkbmalhikmah sukodonoNo ratings yet
- 50 QuestionsDocument2 pages50 QuestionsSzeles GáborNo ratings yet
- Penilaian Akhir Tahun Bahasa Inggeris - Pemahaman (Tahun 3) SK Rancangan SepupokDocument9 pagesPenilaian Akhir Tahun Bahasa Inggeris - Pemahaman (Tahun 3) SK Rancangan SepupokelbrinaNo ratings yet
- Greek Island Ferries Sea Schedules October 09Document72 pagesGreek Island Ferries Sea Schedules October 09SeaSchedulesNo ratings yet
- Nursing AssessmentDocument4 pagesNursing AssessmentFlor SabaysabayNo ratings yet
- Interstitial Vs AlveolarDocument17 pagesInterstitial Vs AlveolarEdwin DarmawanNo ratings yet
- Vedic Recitation and Effects On BrainDocument12 pagesVedic Recitation and Effects On BrainUpadhyayula Surya Raghavender100% (1)
- Roundworm (Ascaris Lumbricoides) : Morphology, Life Cycle & Laboratory DiagnosisDocument14 pagesRoundworm (Ascaris Lumbricoides) : Morphology, Life Cycle & Laboratory DiagnosisUjjawalShriwastavNo ratings yet