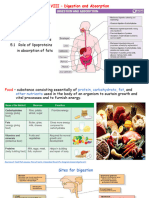
555501661-Carbohydrate Metabolism Bds
555501661-Carbohydrate Metabolism Bds
Download as pdf or txt
You might also like
- Catalogo FMC - Link Belt - Belt Conveyors IdlersDocument124 pagesCatalogo FMC - Link Belt - Belt Conveyors IdlersErich Thomas100% (1)
- Gonstead Tech Study SheetDocument10 pagesGonstead Tech Study Sheetbjpalmer100% (3)
- Venus Uranus Planetary Cycle PDFDocument42 pagesVenus Uranus Planetary Cycle PDFsamaros78% (9)
- Design of Sheet Pile BracingDocument24 pagesDesign of Sheet Pile BracingIsmail Magdy Ismail100% (3)
- Carbohydrate MetabolismDocument139 pagesCarbohydrate MetabolismMarshall AzekeNo ratings yet
- Glycolysis NewDocument45 pagesGlycolysis NewChitra CNo ratings yet
- Chapter 4-IIDocument36 pagesChapter 4-IIefiNo ratings yet
- Carbohydrate MetabolismDocument19 pagesCarbohydrate MetabolismAbhithNo ratings yet
- Digestion and Absorption - 123131Document18 pagesDigestion and Absorption - 123131Johnson UgochukwuNo ratings yet
- Intensive Course 2016 2017 Principles of Biochemistry: Prof Nik Norulaini Nik Ab RahmanDocument73 pagesIntensive Course 2016 2017 Principles of Biochemistry: Prof Nik Norulaini Nik Ab RahmanJayanthi LoganathanNo ratings yet
- 2 - Intro and GlycolysisDocument54 pages2 - Intro and Glycolysiskasozi marvinNo ratings yet
- METABOLISM OF CARBOHYDRATES Part 3Document24 pagesMETABOLISM OF CARBOHYDRATES Part 3akkukesu2002No ratings yet
- InMet I Well-Fed State Spring 23Document32 pagesInMet I Well-Fed State Spring 23Erin HillNo ratings yet
- BIO 202 Biochemistry II by Seyhun YURDUGÜL: Carbohydrate Metabolism I GlycolysisDocument59 pagesBIO 202 Biochemistry II by Seyhun YURDUGÜL: Carbohydrate Metabolism I GlycolysisYasin Çağrı KılıçerNo ratings yet
- Dgestion and Absorption of CHODocument15 pagesDgestion and Absorption of CHOessaNo ratings yet
- DR Samra Hafeez Assistant Professor Dept of BiochemistryDocument54 pagesDR Samra Hafeez Assistant Professor Dept of BiochemistryAttiqaQureshiNo ratings yet
- Muscle Energy Metabolsm For MedicineDocument69 pagesMuscle Energy Metabolsm For MedicinefikaduNo ratings yet
- Glycolysis PDFDocument44 pagesGlycolysis PDFAbdul Jabbar Abdul JabbarNo ratings yet
- Bio Lec1Document32 pagesBio Lec1mernanmmdouh275No ratings yet
- LECTURE 2 - Carbohydrate MetabolismDocument60 pagesLECTURE 2 - Carbohydrate Metabolismmuhammedgmdidra95No ratings yet
- Digestion and Absorption of CarbohydratesDocument25 pagesDigestion and Absorption of CarbohydrateskhadijaNo ratings yet
- CarbohydrateDocument72 pagesCarbohydratedeahajjaj500No ratings yet
- Metabolism Complete Notes #KigogoDocument145 pagesMetabolism Complete Notes #KigogoHesbone AneneNo ratings yet
- NutritionDocument1 pageNutritionUdhaya ShanmugamNo ratings yet
- Course BIO-411Document144 pagesCourse BIO-411Muhammad MoinNo ratings yet
- Carbohydrate MetabolismDocument46 pagesCarbohydrate MetabolismAnahgen RomeoNo ratings yet
- Lec 1 CarbDocument39 pagesLec 1 Carbhendemad719No ratings yet
- 1-Metabolic Fate of Carohydtrates 1Document21 pages1-Metabolic Fate of Carohydtrates 1talhayousaf773No ratings yet
- Digestion and Absorption of CarbohydratesDocument28 pagesDigestion and Absorption of Carbohydratesshakila786No ratings yet
- Integration of Metabolism: Dr. Farzana Hakim Assistant Professor Biochemistry DepartmentDocument63 pagesIntegration of Metabolism: Dr. Farzana Hakim Assistant Professor Biochemistry DepartmentGriffin100% (1)
- CARBOHYDRATE METABOLISM-DrSangitaDocument34 pagesCARBOHYDRATE METABOLISM-DrSangitaA MINOR100% (1)
- 3-4 Carbohydrate Metabolism-1 GlycolysisDocument43 pages3-4 Carbohydrate Metabolism-1 Glycolysis969m9k5ffyNo ratings yet
- Carbohydrate MetabolismDocument101 pagesCarbohydrate Metabolismkjgbitoon00125No ratings yet
- Group Presentation (A) Group Presentation : Health Science (Gen. Nursing) Biochemistry Gluconeogenesis DR SujuudDocument31 pagesGroup Presentation (A) Group Presentation : Health Science (Gen. Nursing) Biochemistry Gluconeogenesis DR SujuudIbrahim WarsameNo ratings yet
- Liver Metabolism: Liver Is The Chief Metabolic Organ of Our BodyDocument123 pagesLiver Metabolism: Liver Is The Chief Metabolic Organ of Our Bodymex Gbrekorkos100% (1)
- Chemistry of CarbohydratesDocument10 pagesChemistry of CarbohydratesYahya Daham Zafeer SakhrNo ratings yet
- CARBOHYDRATE METABOLISM Power Point Lecture NotesDocument32 pagesCARBOHYDRATE METABOLISM Power Point Lecture NotesAnthony AnibasaNo ratings yet
- Lec 3 Biochemistry IDocument34 pagesLec 3 Biochemistry Imaxwellngari728No ratings yet
- ChoDocument44 pagesChosunielgowdaNo ratings yet
- Lecture 03. Carbohydrate Metabolism. Common Pathway, Regulation and PathologyDocument80 pagesLecture 03. Carbohydrate Metabolism. Common Pathway, Regulation and PathologyВіталій Михайлович НечипорукNo ratings yet
- Metabolisme Dan TermoregulasiDocument56 pagesMetabolisme Dan TermoregulasiMuhammad Iqbal100% (1)
- Biochemistry Notes 2Document903 pagesBiochemistry Notes 2madhu PriyaNo ratings yet
- GlycolysisDocument17 pagesGlycolysisSourav DasNo ratings yet
- Lecture 02 (Function of GI Tract in Digestion and Adsorption)Document45 pagesLecture 02 (Function of GI Tract in Digestion and Adsorption)عبدالرحمن دودينNo ratings yet
- Unit 8 Digestion and AbsorptionDocument33 pagesUnit 8 Digestion and AbsorptionGlory MuedaNo ratings yet
- CarbohydratemetabolismDocument88 pagesCarbohydratemetabolismEthio funNo ratings yet
- Carbohydrates Metabolism Lecture 2021Document120 pagesCarbohydrates Metabolism Lecture 2021Manish ShahNo ratings yet
- Sehs Topic 3 2 Carbohydrate and Fat MetabolismDocument20 pagesSehs Topic 3 2 Carbohydrate and Fat Metabolismapi-343368893No ratings yet
- Digestion and Absorption of CarbohydratesDocument17 pagesDigestion and Absorption of Carbohydratesm.ibrahim.dhilavarNo ratings yet
- Glyconeogenesis - BIO 3110Document23 pagesGlyconeogenesis - BIO 3110NaiomiNo ratings yet
- Metabolism of Carbohydrates-1Document64 pagesMetabolism of Carbohydrates-1Amina AmiNo ratings yet
- Gluconeogenesis NewDocument10 pagesGluconeogenesis Newaqsasiddiqui337No ratings yet
- 01 Regulation of Organic Metabolism and Energy BalanceDocument65 pages01 Regulation of Organic Metabolism and Energy Balancetaserullah95No ratings yet
- Metabolisme KarbohidratDocument56 pagesMetabolisme KarbohidratAnonymous QCMhA4wNgBNo ratings yet
- Digestion and AbsorptionDocument10 pagesDigestion and AbsorptionsilentdestructionNo ratings yet
- Carbohydrates Part 2 - MetabolismsDocument60 pagesCarbohydrates Part 2 - MetabolismsNikka Mia AbadiesNo ratings yet
- Week 6 LipidsDocument59 pagesWeek 6 Lipidsbo aNo ratings yet
- Glycolysis 1Document33 pagesGlycolysis 1AhmedA.SharafNo ratings yet
- Integration of MetabolismDocument17 pagesIntegration of MetabolismSharif M Mizanur RahmanNo ratings yet
- 3.2. Carbo-Meta - Short VersionDocument29 pages3.2. Carbo-Meta - Short VersionLong NguyenNo ratings yet
- Specific Catabolic PathwaysDocument22 pagesSpecific Catabolic Pathwaysmimingk.andraNo ratings yet
- Physics ProjectDocument12 pagesPhysics ProjectDanish SharmaNo ratings yet
- Revision Notes On Co-Ordination CompoundsDocument12 pagesRevision Notes On Co-Ordination CompoundsAnonymous vRpzQ2BLNo ratings yet
- EGCSE Mathematics 2024-2026 SyllabusDocument31 pagesEGCSE Mathematics 2024-2026 Syllabussena06.mathenjwaNo ratings yet
- Quiz#3 1Document3 pagesQuiz#3 1Ronalyn AsunioNo ratings yet
- Wastewater and Biosolids Analysis Manual PDFDocument512 pagesWastewater and Biosolids Analysis Manual PDFTuyen NguyenNo ratings yet
- 2 Sabol Bim FacilityDocument13 pages2 Sabol Bim FacilityTareq Ali طارق عليNo ratings yet
- An Overview of Device-To-Device Communication in Cellular NetworksDocument6 pagesAn Overview of Device-To-Device Communication in Cellular NetworksZiya2009No ratings yet
- WISENT Cidds Technical ReportDocument8 pagesWISENT Cidds Technical ReportsantiNo ratings yet
- Diesel Engine Fundamentals HandbookDocument236 pagesDiesel Engine Fundamentals HandbookMiguel Camacho100% (2)
- Module 2c - NR RecapDocument8 pagesModule 2c - NR RecapChen HaoNo ratings yet
- Rubiks-Cube 3x3 PLLDocument4 pagesRubiks-Cube 3x3 PLLHarrey MaurealNo ratings yet
- Old Gui With SwingDocument319 pagesOld Gui With SwingPajith PajithNo ratings yet
- Mathletics Grade 3 - FractionsDocument25 pagesMathletics Grade 3 - FractionsMadeleine EsperanzaNo ratings yet
- 9tech Spec 100 To 2500 KVA Dry Type Dist - Tranformer 1Document30 pages9tech Spec 100 To 2500 KVA Dry Type Dist - Tranformer 1Yazhisai SelviNo ratings yet
- GS 142-2Document31 pagesGS 142-2Serge RINAUDO0% (1)
- Chapter 1 - Intro To Programming Language 20182019Document54 pagesChapter 1 - Intro To Programming Language 20182019Abdul Ashraf RasidNo ratings yet
- Sixto Ramos Fernando SolanoDocument17 pagesSixto Ramos Fernando SolanoMichael PrzybylskiNo ratings yet
- Reference For MooringDocument20 pagesReference For MooringPeyman MznNo ratings yet
- AThakkar QChem BookDocument122 pagesAThakkar QChem BookMahmoud SalemNo ratings yet
- Manual Controlador Factor Potencia 320Document15 pagesManual Controlador Factor Potencia 320Hector_Cornejo_5673No ratings yet
- Chapter 3 Cascaded Style Sheets (CSS)Document21 pagesChapter 3 Cascaded Style Sheets (CSS)zeki.mama21.21No ratings yet
- SML Technology Report 2. Edition 2010 Druckversion v. 12.10.10 SaveDocument0 pagesSML Technology Report 2. Edition 2010 Druckversion v. 12.10.10 SaveMarcWorldNo ratings yet
- 13 - Finishing EquipmentDocument9 pages13 - Finishing EquipmentKArthik AustinNo ratings yet
- Advanced Drum LessonsDocument26 pagesAdvanced Drum LessonsBruno Gonçalves100% (1)
- Ain Shams Engineering Journal: Engineering Physics and MathematicsDocument15 pagesAin Shams Engineering Journal: Engineering Physics and MathematicsMbarouk Shaame MbaroukNo ratings yet
- Gantrail Welded Base Fixing: SpecificationsDocument2 pagesGantrail Welded Base Fixing: SpecificationsCarlos Augusto LancellottiNo ratings yet