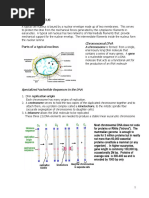
DNA Topology and Supercoiling
DNA Topology and Supercoiling
Download as pptx, pdf, or txt
You might also like
- MBY 364 NOTES - MergedDocument22 pagesMBY 364 NOTES - Mergedroelienvv7No ratings yet
- Production and Operations Improvement Plan (POIP) : Manufaturing Business (Corporation Type)Document30 pagesProduction and Operations Improvement Plan (POIP) : Manufaturing Business (Corporation Type)nikNo ratings yet
- BTM104-Lecture4 2022Document35 pagesBTM104-Lecture4 2022ginirajput12No ratings yet
- Lecture 2 - DNA & ChromosomeDocument74 pagesLecture 2 - DNA & ChromosomeConstance WongNo ratings yet
- DNA SupercoilingDocument33 pagesDNA SupercoilingPRANATHI REDDYNo ratings yet
- MBD LessonsDocument9 pagesMBD LessonsAnna Charisa HernandezNo ratings yet
- BTN222-L2-DNA and Genome STR-2019Document43 pagesBTN222-L2-DNA and Genome STR-2019rafiya IsmailNo ratings yet
- Chromosomes and DNA Packaging - LectureDocument34 pagesChromosomes and DNA Packaging - LectureNicol AndreescuNo ratings yet
- DNA Estructura, TopologíaDocument11 pagesDNA Estructura, TopologíaMariana Sandoval NosetyNo ratings yet
- BMB801 Fall 2020 Lecture Period 4: DNA TopologyDocument13 pagesBMB801 Fall 2020 Lecture Period 4: DNA TopologyIda ShafieiNo ratings yet
- Lec-1 (24-09-2024)Document10 pagesLec-1 (24-09-2024)Kritika ShreeNo ratings yet
- MCB 110 Study GuideDocument10 pagesMCB 110 Study GuideAlexPowersNo ratings yet
- Functions of The Genetic MaterialDocument31 pagesFunctions of The Genetic MaterialZara HussainNo ratings yet
- Genetics Seminars 1-3Document289 pagesGenetics Seminars 1-3Мөнхгэрэл ГанбатNo ratings yet
- Lecture 1-Genetic Material, DNA Structure and Replication-2Document46 pagesLecture 1-Genetic Material, DNA Structure and Replication-2UJPNo ratings yet
- Unit 2Document59 pagesUnit 2mishtydprNo ratings yet
- cc8t DnaDocument25 pagescc8t DnaRiya SinghNo ratings yet
- ٣٢٥ Molecular Biology Sabah Linjawi ١Document16 pages٣٢٥ Molecular Biology Sabah Linjawi ١Zainab RaikNo ratings yet
- Genes Are Dna and Encode Rnas and PolypeptidesDocument66 pagesGenes Are Dna and Encode Rnas and Polypeptidesshaurya singhNo ratings yet
- SMIB032 Lecture #1 15 07 2024Document31 pagesSMIB032 Lecture #1 15 07 2024lion2ndtNo ratings yet
- Mod 2 - The NucleusDocument5 pagesMod 2 - The Nucleusaljunma04No ratings yet
- ReplicationDocument58 pagesReplicationBillie LunaNo ratings yet
- Lesson 2. Nucleus and chromosomes . DesyatovaDocument40 pagesLesson 2. Nucleus and chromosomes . Desyatovahimanshisaini0904No ratings yet
- BiochemistryDocument48 pagesBiochemistrylittle wordsNo ratings yet
- Bio Ninja Unit 7 MergedDocument4 pagesBio Ninja Unit 7 Mergedshwooby shawoobyNo ratings yet
- Bacterial GeneticsDocument38 pagesBacterial Geneticsfatima zafarNo ratings yet
- DNA PackagingDocument25 pagesDNA PackagingChamara ChathurangaNo ratings yet
- DNA ReplicationDocument12 pagesDNA Replicationlaserdog97No ratings yet
- Topic 7 Nucleic AcidsDocument19 pagesTopic 7 Nucleic AcidsSammy MicahNo ratings yet
- Chromosome Organization & DNA Packaging: BME 42-620 Engineering Molecular Cell BiologyDocument35 pagesChromosome Organization & DNA Packaging: BME 42-620 Engineering Molecular Cell BiologyNermeen ElmelegaeNo ratings yet
- DNA Packaging by Enamul HaqueDocument27 pagesDNA Packaging by Enamul HaqueEnamul Haque100% (1)
- Dnareplication 151218084840Document145 pagesDnareplication 151218084840RIYA MUNJALNo ratings yet
- Prokaryotic Replication a Journey Inside the CellDocument10 pagesProkaryotic Replication a Journey Inside the Cellzohebqaiser02No ratings yet
- MICROBIO 1.3 Bacterial GeneticsDocument6 pagesMICROBIO 1.3 Bacterial GeneticsPatricia Elena ManaliliNo ratings yet
- Lehninger Principles of Biochemistry: Fourth EditionDocument105 pagesLehninger Principles of Biochemistry: Fourth EditionFahad JavedNo ratings yet
- Materi Minggu 6Document44 pagesMateri Minggu 6HafizahNo ratings yet
- 1 DNA Structure and ReplicationDocument96 pages1 DNA Structure and ReplicationmattMd100% (1)
- Unit 3. The NucleusDocument130 pagesUnit 3. The NucleusArio AhmadieNo ratings yet
- Dna, Genes and GenomesDocument4 pagesDna, Genes and GenomesRobert Martin PuertaNo ratings yet
- Molecular Genetics Chapter 8Document49 pagesMolecular Genetics Chapter 8byunus88No ratings yet
- Histone Proteins, The Nucleosome and Chromatin Structure - 9Document44 pagesHistone Proteins, The Nucleosome and Chromatin Structure - 9REMYAREMA31No ratings yet
- Summary OCPEG and Cancer 2018Document18 pagesSummary OCPEG and Cancer 2018Jonathan Wee Cheng YangNo ratings yet
- Genome StructureDocument62 pagesGenome Structuregeet maheshwariNo ratings yet
- The Cell Nucleus: Nuclear LaminaDocument5 pagesThe Cell Nucleus: Nuclear LaminaRealicahrealicah RealicahrealicahNo ratings yet
- Lecture Notes 2 (Central Dogma)Document9 pagesLecture Notes 2 (Central Dogma)Lorraine M. Del RosarioNo ratings yet
- Molecular Genetics Chapter 3Document5 pagesMolecular Genetics Chapter 3api-508390609No ratings yet
- Molecular BiologyDocument52 pagesMolecular Biologychandu SahaNo ratings yet
- Lecture 3 (06) WebsiteDocument27 pagesLecture 3 (06) Websiteeri512457No ratings yet
- Chapter 8 Cell ReproductionDocument20 pagesChapter 8 Cell ReproductionDaniel ChrysNo ratings yet
- MB Chapter 4 - DNA-Replication-IDocument27 pagesMB Chapter 4 - DNA-Replication-IMustee TeferaNo ratings yet
- Prokaryotic DNA ReplicationDocument31 pagesProkaryotic DNA ReplicationAnonymous Ef3jslp8No ratings yet
- DNA RepairDocument98 pagesDNA Repairapi-3700537100% (1)
- Chemical Basis of HeredityDocument21 pagesChemical Basis of HeredityEmelinda CruzNo ratings yet
- ChromatinDocument35 pagesChromatinKhushnood FatimaNo ratings yet
- Hóa Sinh Phân T 1Document13 pagesHóa Sinh Phân T 1Quang Huỳnh ViệtNo ratings yet
- Dna ReplicationDocument145 pagesDna ReplicationDrMumtaz F MusaliarNo ratings yet
- Chromosome, DNA, RNADocument5 pagesChromosome, DNA, RNAShahrat Farsim ChowdhuryNo ratings yet
- Lehninger PPT Ch24Document61 pagesLehninger PPT Ch24Nia IarajuliNo ratings yet
- 1 Los Genes Son ADNDocument24 pages1 Los Genes Son ADNEmanuel Rodriguez ArrazateNo ratings yet
- 2.chemistry of Nucleic AcidsDocument46 pages2.chemistry of Nucleic AcidsYitbarek Nigat AlemuNo ratings yet
- Installation of Inductive Loop Detectors: Standard Practice For TheDocument13 pagesInstallation of Inductive Loop Detectors: Standard Practice For ThearaNo ratings yet
- Criminal Complaint State of Wisconsin v. Armor Correctional Health Services, Inc.Document11 pagesCriminal Complaint State of Wisconsin v. Armor Correctional Health Services, Inc.CHRISTINA CURRIENo ratings yet
- Organic Agriculture GlossaryDocument65 pagesOrganic Agriculture GlossaryAhmed Abd ElNasser RabieNo ratings yet
- Trane Product PL Rev 6-1-2023Document20 pagesTrane Product PL Rev 6-1-2023edizciperNo ratings yet
- Ore-Forming Fluid Source of The Orogenic Gold DepositDocument57 pagesOre-Forming Fluid Source of The Orogenic Gold DepositHeri GeologistNo ratings yet
- Evidence Based Practice in Educating Deaf and Hard of Hearing Students Professional Perspectives on Deafness Evidence and Applications 1st Edition Patricia Elizabeth Spencer All Chapters Instant DownloadDocument82 pagesEvidence Based Practice in Educating Deaf and Hard of Hearing Students Professional Perspectives on Deafness Evidence and Applications 1st Edition Patricia Elizabeth Spencer All Chapters Instant Downloadalmissshamsa100% (8)
- 6 Big Losses in Manufacturing-2Document3 pages6 Big Losses in Manufacturing-2izzudinrozNo ratings yet
- Simple ReceiptDocument4 pagesSimple ReceiptMark AbraganNo ratings yet
- Thermography Evaluation of Low Back Pain in Pregnant Women Cross-Sectional StudyDocument5 pagesThermography Evaluation of Low Back Pain in Pregnant Women Cross-Sectional Studymayco.andersonNo ratings yet
- OH Crane DemagDocument13 pagesOH Crane DemagTeJeS farioNo ratings yet
- Streptococcus PyogenesDocument40 pagesStreptococcus Pyogenesnshree155No ratings yet
- Vandana Raut Tata Memorial Centre 31yrsDocument2 pagesVandana Raut Tata Memorial Centre 31yrsvandanaNo ratings yet
- Good Grounding PracticesDocument24 pagesGood Grounding PracticesSamia NazNo ratings yet
- TSS - Copper Tube - REV5Document8 pagesTSS - Copper Tube - REV5Kaleem MaharNo ratings yet
- Bio-Mass Gasification: 15Eee359-Renewable Energy and Energy ConservationDocument13 pagesBio-Mass Gasification: 15Eee359-Renewable Energy and Energy ConservationKarthikeyan GNo ratings yet
- Dealogic DCM CriteriaDocument3 pagesDealogic DCM Criteriavineet_bmNo ratings yet
- Small Business FSMS Implementation Guide SampleDocument32 pagesSmall Business FSMS Implementation Guide SampleGaganpreet KaurNo ratings yet
- Datasheet Leapton Panther 570 LP182 199 M 66 NHDocument2 pagesDatasheet Leapton Panther 570 LP182 199 M 66 NHrodrigoNo ratings yet
- Dual Pressure Module, E-PP Pressure Temp Module, E-PT: Service ManualDocument46 pagesDual Pressure Module, E-PP Pressure Temp Module, E-PT: Service Manualjhon aguileraNo ratings yet
- Accord Product ListDocument14 pagesAccord Product ListANIRUDDHA KAPADNISNo ratings yet
- Peritoneal DialysisDocument12 pagesPeritoneal DialysisSania TohattaNo ratings yet
- Plant NutrientsDocument3 pagesPlant NutrientsAbdulrehman NizamaniNo ratings yet
- AEROSOL OT100% Surf Act Ant (USA Version)Document6 pagesAEROSOL OT100% Surf Act Ant (USA Version)kohonNo ratings yet
- March 2010 Lighthouse Point MagazineDocument76 pagesMarch 2010 Lighthouse Point MagazineJon FrangipaneNo ratings yet
- The Other Half of NormalDocument118 pagesThe Other Half of NormalJen Gossman ColemanNo ratings yet
- 1622-DES-CR-02 - Gas Cylinder Basket Design Report - R0Document23 pages1622-DES-CR-02 - Gas Cylinder Basket Design Report - R0Aditya Jain100% (1)
- Kamus PANDocument77 pagesKamus PANserdarcico1209No ratings yet
- Bene Bay Chair Data SheetDocument5 pagesBene Bay Chair Data SheetcincinikNo ratings yet
- Practical Religion by Swami VivekanandDocument5 pagesPractical Religion by Swami VivekanandTanisha sighalNo ratings yet