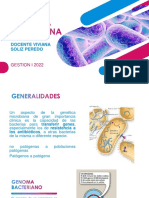
Conjugación Procariota
Conjugación Procariota
Descargar como doc, pdf o txt
También podría gustarte
- Elementos Genéticos MóvilesDocumento41 páginasElementos Genéticos Móvilesmajo gs100% (2)
- Genetica MicrobianaDocumento71 páginasGenetica MicrobianaKNayAún no hay calificaciones
- FlagelosDocumento18 páginasFlagelosJovan Rivero MeralesAún no hay calificaciones
- ConjugacionDocumento3 páginasConjugacionOscar JimenezAún no hay calificaciones
- Marco Teorico de MicroDocumento3 páginasMarco Teorico de MicroMimi MartinezAún no hay calificaciones
- Taller N°2 Biologia MolecularDocumento9 páginasTaller N°2 Biologia Molecularrobinalbarracin3Aún no hay calificaciones
- Microbiología - Taller PDFDocumento6 páginasMicrobiología - Taller PDFvaluAún no hay calificaciones
- 01 - REV02 - Grupo01 - Seminario 1.1. Conjugación Procariota, Transformación y Transducción - Sem3Documento13 páginas01 - REV02 - Grupo01 - Seminario 1.1. Conjugación Procariota, Transformación y Transducción - Sem3guesorrock 888Aún no hay calificaciones
- Control Genético en ProcariotasDocumento9 páginasControl Genético en ProcariotasleomelthormeAún no hay calificaciones
- Genetica Bacteriana y Patogenicidad Bacteriana ResumenDocumento4 páginasGenetica Bacteriana y Patogenicidad Bacteriana ResumenMarina ArmentaAún no hay calificaciones
- Trabajo No.16Documento5 páginasTrabajo No.16SERGIO DIAZ VILLANUEVAAún no hay calificaciones
- Plasmidos de MolecularDocumento7 páginasPlasmidos de Molecularandres zuluagaAún no hay calificaciones
- Clase 7Documento19 páginasClase 7MaximimilianoAún no hay calificaciones
- CAPÍTULO 8 (Guía de TP 2024) RECOMBINACIÓN EN BACTERIAS-AVDocumento6 páginasCAPÍTULO 8 (Guía de TP 2024) RECOMBINACIÓN EN BACTERIAS-AVayelen.cuello.dominguezAún no hay calificaciones
- Mecanísmos de Variación GenéticaDocumento5 páginasMecanísmos de Variación GenéticaCatalina Duque HenaoAún no hay calificaciones
- Clase HGT-ConjugaciónDocumento58 páginasClase HGT-ConjugaciónPortilla AnngyAún no hay calificaciones
- Genoma ProcariotaDocumento14 páginasGenoma ProcariotaAbel MooreAún no hay calificaciones
- Regulación Genética en ProcariotasDocumento15 páginasRegulación Genética en ProcariotasjelAún no hay calificaciones
- Resumen Genetica Parte 2Documento38 páginasResumen Genetica Parte 2CAMIAún no hay calificaciones
- Genética BacterianaDocumento29 páginasGenética BacterianaLuisAún no hay calificaciones
- ConjugacionDocumento3 páginasConjugacionsandra osornioAún no hay calificaciones
- El Tipo de ReproducciónDocumento3 páginasEl Tipo de ReproducciónSOFI FERNANDEZAún no hay calificaciones
- La Reproduccion BacterianaDocumento4 páginasLa Reproduccion BacterianayamilkaAún no hay calificaciones
- Resumen 5 MicroDocumento11 páginasResumen 5 Microhv66580Aún no hay calificaciones
- Micro Biolog I ADocumento7 páginasMicro Biolog I AAnthony LaderaAún no hay calificaciones
- Recombinacion Bacteriana Documento TrabajoDocumento33 páginasRecombinacion Bacteriana Documento TrabajoWillSmith SanchezAún no hay calificaciones
- 23 (28-04) ConjugaciónDocumento8 páginas23 (28-04) ConjugaciónValeria MoragaAún no hay calificaciones
- Recombinación BacterianaDocumento23 páginasRecombinación BacterianaStefany Beni Ruiz50% (2)
- Capítulo 16 - Recombinación en BacteriasDocumento8 páginasCapítulo 16 - Recombinación en Bacteriasdiego ascuntarAún no hay calificaciones
- Transferencia de Genes FinalDocumento22 páginasTransferencia de Genes FinalMelisa Quimiz OrtegaAún no hay calificaciones
- Conjugacion 4Documento31 páginasConjugacion 4Magdiel Cruz OliveraAún no hay calificaciones
- Genetica Bacteriana 2022Documento51 páginasGenetica Bacteriana 2022Bryan 015Aún no hay calificaciones
- Reproducción en Las BacteriasDocumento7 páginasReproducción en Las BacteriasJose AyarzaAún no hay calificaciones
- Resumen A1Documento1 páginaResumen A1karlajvgarcia.03Aún no hay calificaciones
- TransposonesDocumento4 páginasTransposonesJessica CruzAún no hay calificaciones
- Taller Genetica BacterianaDocumento8 páginasTaller Genetica BacterianalinaAún no hay calificaciones
- Tema 2 Genetica Bacteriana #2Documento33 páginasTema 2 Genetica Bacteriana #2Juan ChambiAún no hay calificaciones
- Recombinación Genética PDFDocumento5 páginasRecombinación Genética PDFJavan ZaldivarAún no hay calificaciones
- Recombinacion Genetica BacterianaDocumento37 páginasRecombinacion Genetica Bacterianajesus david Alvarez truyolAún no hay calificaciones
- Elementos Geneticos de Resistencia en La Poblacion MicrobianaDocumento20 páginasElementos Geneticos de Resistencia en La Poblacion MicrobianaSamantha LopezAún no hay calificaciones
- Genoma BacterianoDocumento4 páginasGenoma BacterianoMiguel MacdonaldAún no hay calificaciones
- Capitulo 7 JawetzDocumento5 páginasCapitulo 7 Jawetzy57h9tnt2dAún no hay calificaciones
- Transferencia de Material GeneticoDocumento22 páginasTransferencia de Material GeneticoGabriela BuendiaAún no hay calificaciones
- TransposónDocumento7 páginasTransposónAntonio TreminiAún no hay calificaciones
- PlasmidosDocumento62 páginasPlasmidosBrandon Chahua ObregonAún no hay calificaciones
- Recombinacion GeneticaDocumento3 páginasRecombinacion GeneticaAntony SIESQUEN DIAZAún no hay calificaciones
- Multiplicacion de La Celula BacterianaDocumento7 páginasMultiplicacion de La Celula BacterianaMaría Lucero HobaicaAún no hay calificaciones
- Crecimiento, Reproducción y Mutación BacterianoDocumento24 páginasCrecimiento, Reproducción y Mutación BacterianoJair Benavente AsinAún no hay calificaciones
- Recombinación Especifica de Sitio y TransposicionalDocumento3 páginasRecombinación Especifica de Sitio y TransposicionalBibiana VarónAún no hay calificaciones
- Genetica BacterianaDocumento22 páginasGenetica Bacterianaaxel_gutierrez_11Aún no hay calificaciones
- 10 RecombinacionDocumento5 páginas10 RecombinacionankylosaurioAún no hay calificaciones
- Genetica BacterianaDocumento24 páginasGenetica Bacteriana8r9vns72rfAún no hay calificaciones
- Conjugación TGH: Fisiología BacterianaDocumento4 páginasConjugación TGH: Fisiología BacterianaMarina ChaconAún no hay calificaciones
- Cuestionario-Genética BCDocumento5 páginasCuestionario-Genética BCKarlo MartinesAún no hay calificaciones
- Microbiologia Presentación Power PointDocumento25 páginasMicrobiologia Presentación Power PointGil GopAún no hay calificaciones
- Genetica MicrobianaDocumento20 páginasGenetica MicrobianaWilman Manuel Benavidez HernándezAún no hay calificaciones
- Recombinación Heteróloga - BFDocumento9 páginasRecombinación Heteróloga - BFPaula Estefania Urrego VillalobosAún no hay calificaciones
- Genética Bacteriana y Ecología BacterianaDocumento13 páginasGenética Bacteriana y Ecología BacterianaValeria VAún no hay calificaciones
- S 2a Semana 2 a Clonación y transgénesis en procariotas, células vegetales.Documento11 páginasS 2a Semana 2 a Clonación y transgénesis en procariotas, células vegetales.Joe Dangelo GSAún no hay calificaciones
- Genetica Bacteriana 129955 Downloable 1905806Documento5 páginasGenetica Bacteriana 129955 Downloable 1905806Daniela Garcia CondeAún no hay calificaciones
- Genética MicrobianaDocumento3 páginasGenética MicrobianaKeyla VegaAún no hay calificaciones
- Período de OscilaciónDocumento4 páginasPeríodo de OscilaciónAntonio TreminiAún no hay calificaciones
- GigahercioDocumento2 páginasGigahercioAntonio TreminiAún no hay calificaciones
- Geología de CaronteDocumento2 páginasGeología de CaronteAntonio TreminiAún no hay calificaciones
- MicroondasDocumento9 páginasMicroondasAntonio TreminiAún no hay calificaciones
- New Horizons (Sonda Espacial)Documento7 páginasNew Horizons (Sonda Espacial)Antonio TreminiAún no hay calificaciones
- Caronte (Satélite)Documento5 páginasCaronte (Satélite)Antonio TreminiAún no hay calificaciones
- PlutoideDocumento2 páginasPlutoideAntonio TreminiAún no hay calificaciones
- Haumea (Planeta Enano)Documento4 páginasHaumea (Planeta Enano)Antonio TreminiAún no hay calificaciones
- Makemake (Planeta Enano)Documento4 páginasMakemake (Planeta Enano)Antonio TreminiAún no hay calificaciones
- Objeto TransneptunianoDocumento2 páginasObjeto TransneptunianoAntonio TreminiAún no hay calificaciones
- PlanetesimalDocumento3 páginasPlanetesimalAntonio TreminiAún no hay calificaciones
- (50000) QuaoarDocumento4 páginas(50000) QuaoarAntonio TreminiAún no hay calificaciones
- (15760) AlbionDocumento3 páginas(15760) AlbionAntonio TreminiAún no hay calificaciones
- (486958) ArrokothDocumento4 páginas(486958) ArrokothAntonio TreminiAún no hay calificaciones
- (20000) VarunaDocumento4 páginas(20000) VarunaAntonio TreminiAún no hay calificaciones
- PlanetoideDocumento2 páginasPlanetoideAntonio TreminiAún no hay calificaciones
- Cometa ExtintoDocumento1 páginaCometa ExtintoAntonio TreminiAún no hay calificaciones
- Cometa HalleyDocumento3 páginasCometa HalleyAntonio TreminiAún no hay calificaciones
- Asteroide AtónDocumento1 páginaAsteroide AtónAntonio TreminiAún no hay calificaciones
- Churyumov GerasimenkoDocumento4 páginasChuryumov GerasimenkoAntonio TreminiAún no hay calificaciones
- BorrellyDocumento3 páginasBorrellyAntonio TreminiAún no hay calificaciones
- Asteroide AmorDocumento2 páginasAsteroide AmorAntonio TreminiAún no hay calificaciones
- (1566) IcarusDocumento3 páginas(1566) IcarusAntonio TreminiAún no hay calificaciones
- (2063) BacchusDocumento3 páginas(2063) BacchusAntonio TreminiAún no hay calificaciones
- (1866) SisyphusDocumento3 páginas(1866) SisyphusAntonio TreminiAún no hay calificaciones
- Guía de Actividades y Rúbrica de Evaluación - Tarea 1 - Microorganismos ProcariotasDocumento9 páginasGuía de Actividades y Rúbrica de Evaluación - Tarea 1 - Microorganismos ProcariotaslilianaAún no hay calificaciones
- Tema 33: Reino Moneras. Las Cyanophytas. Las Bacterias Y Su Importancia en La Sanidad, La Industria Y La Investigacion Básica 1. IntroducciónDocumento13 páginasTema 33: Reino Moneras. Las Cyanophytas. Las Bacterias Y Su Importancia en La Sanidad, La Industria Y La Investigacion Básica 1. IntroducciónPaco BejaranoAún no hay calificaciones
- Tema 1: Introducción A La Genética GenéticaDocumento167 páginasTema 1: Introducción A La Genética GenéticaMelisa BarbasAún no hay calificaciones
- Genetica Bacteriana - AplicacionesDocumento41 páginasGenetica Bacteriana - AplicacionesDayanaAún no hay calificaciones
- Mecanismos de Evolución y Resistencia BacterianasDocumento4 páginasMecanismos de Evolución y Resistencia BacterianasValeria B. MuñincoAún no hay calificaciones
- Examen Modulo 3Documento7 páginasExamen Modulo 3mónica bravo valleAún no hay calificaciones
- Guía Primer Parcial de Introducción A La GenéticaDocumento5 páginasGuía Primer Parcial de Introducción A La GenéticaNorberto Singh Rios (Norbit)Aún no hay calificaciones
- PlasmidosDocumento11 páginasPlasmidosanon_949478666Aún no hay calificaciones
- 01 MicrobiologiaDocumento113 páginas01 Microbiologiajabonete100% (2)
- Organizacion Celular PDFDocumento42 páginasOrganizacion Celular PDFProfaRebecaManriqueAún no hay calificaciones
- Transferencia Del Material Genetico en BacteriasDocumento14 páginasTransferencia Del Material Genetico en BacteriasEdinson Anillo ArrietaAún no hay calificaciones
- GTP - T 12.células (2 Parte - Células Procariotas) 2013-15Documento16 páginasGTP - T 12.células (2 Parte - Células Procariotas) 2013-15Franklin garryAún no hay calificaciones
- Manual AdixDocumento31 páginasManual AdixjoseberrioAún no hay calificaciones
- Celula ProcariotaDocumento8 páginasCelula ProcariotaDany MoralesAún no hay calificaciones
- Microbiologia II UNIDAD II Alejandra Alvarado - Samuel ReyesDocumento62 páginasMicrobiologia II UNIDAD II Alejandra Alvarado - Samuel ReyesBrando Teran100% (3)
- Guía de Antecedentes para Preparación de Seminarios Del Laboratorio de GenéticaDocumento3 páginasGuía de Antecedentes para Preparación de Seminarios Del Laboratorio de GenéticaTania Karen Rodriguez GarciaAún no hay calificaciones
- La Recombinación Del ADNDocumento7 páginasLa Recombinación Del ADNYahir AriasAún no hay calificaciones
- Reproducción BacterianaDocumento17 páginasReproducción BacterianaOriana Hnnd'z-mAún no hay calificaciones
- Biología RecombinacionDocumento3 páginasBiología RecombinacionCesarAún no hay calificaciones
- Micro LeticiaDocumento48 páginasMicro LeticiaCRISTOBAL EDUARDO ALBORNOZ PEREZAún no hay calificaciones
- Micro Falta ResolverDocumento16 páginasMicro Falta ResolverKevin AndresAún no hay calificaciones
- Infect oDocumento89 páginasInfect oDaniel ChávezAún no hay calificaciones
- MICROBIOLOGIADocumento58 páginasMICROBIOLOGIAmarco100% (1)
- Guía de Genética BacterianaDocumento4 páginasGuía de Genética BacterianaJl ReyesAún no hay calificaciones
- Guía de Actividades y Rúbrica de Evaluación - Tarea 1 - Microorganismos ProcariotasDocumento10 páginasGuía de Actividades y Rúbrica de Evaluación - Tarea 1 - Microorganismos ProcariotasERiKA REYESAún no hay calificaciones
- UntitledDocumento64 páginasUntitledAndres Esteban Roman MatteucciAún no hay calificaciones
- BACTERIASDocumento17 páginasBACTERIASDiana Kareen100% (1)