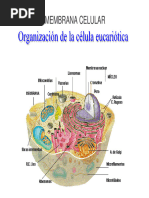
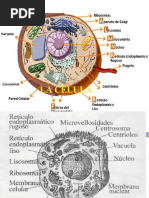
22avo Teo TRANSITO VESICULAR 1
22avo Teo TRANSITO VESICULAR 1
Descargar como pdf o txt
También podría gustarte
- Resumen BCM HistologiaDocumento31 páginasResumen BCM HistologiaJuan Brun50% (2)
- Guia Area de Salud 2022-1Documento80 páginasGuia Area de Salud 2022-1Karina Mg100% (1)
- 24-25 Tráfico Vesicular 1y2Documento9 páginas24-25 Tráfico Vesicular 1y2rosariolema55Aún no hay calificaciones
- Transporte VesicularDocumento6 páginasTransporte VesicularAndrésAún no hay calificaciones
- Actividad 6Documento3 páginasActividad 6Yoselin PioAún no hay calificaciones
- EndosomasDocumento3 páginasEndosomasAriana ReyesAún no hay calificaciones
- SEMDocumento11 páginasSEMjulissagonzalezzuritaAún no hay calificaciones
- Complejos de Union y Transporte Vesicular.Documento12 páginasComplejos de Union y Transporte Vesicular.samiragomez0502Aún no hay calificaciones
- Modulo III Reticulo Endoplásmicosistema de Endomembranas Con Audio Final 2023Documento29 páginasModulo III Reticulo Endoplásmicosistema de Endomembranas Con Audio Final 2023Madelin BolañosAún no hay calificaciones
- Wuolah Free TEMA 5. TRANSITO VESICULARDocumento7 páginasWuolah Free TEMA 5. TRANSITO VESICULARnere.cisferAún no hay calificaciones
- Compartimentos IntracelularesDocumento32 páginasCompartimentos IntracelularesJimena BustamanteAún no hay calificaciones
- Tráfico VesicularDocumento55 páginasTráfico VesicularAficionadoAún no hay calificaciones
- 02 Membrana CitoplasmaticaDocumento73 páginas02 Membrana CitoplasmaticahhgggAún no hay calificaciones
- Biologia SEMANA 9Documento4 páginasBiologia SEMANA 9john salchichon67% (9)
- Aparato de GolgiDocumento12 páginasAparato de Golgial056123028Aún no hay calificaciones
- Microbiologia 13Documento9 páginasMicrobiologia 13David GuidoAún no hay calificaciones
- Bio Cel 3 - Membranas CelularesDocumento9 páginasBio Cel 3 - Membranas CelularesCande FierroAún no hay calificaciones
- Biologia Celular Banco de Preguntas FinalDocumento14 páginasBiologia Celular Banco de Preguntas Finalcharit martinez ñomaAún no hay calificaciones
- Estructura de La Membrana y TransporteDocumento15 páginasEstructura de La Membrana y TransporteAntonieta OtárolaAún no hay calificaciones
- Biocel Solemne 2Documento37 páginasBiocel Solemne 2Keira GonzalezAún no hay calificaciones
- RESUMEN BIOCEL 2.3 Sistema EndomembranasDocumento6 páginasRESUMEN BIOCEL 2.3 Sistema EndomembranasAgustina IribarrenAún no hay calificaciones
- Membrana PlasmaticaDocumento18 páginasMembrana PlasmaticaBorsalinoAún no hay calificaciones
- TEMA5,2 BiocelDocumento18 páginasTEMA5,2 BiocelMóni GonzálezAún no hay calificaciones
- Membrana CelularDocumento28 páginasMembrana CelularAdriana Arianny Diaz CastañedaAún no hay calificaciones
- Sistema de EndomembranasDocumento9 páginasSistema de EndomembranasBelen IñigoAún no hay calificaciones
- Sistema de Endomembranas IDocumento9 páginasSistema de Endomembranas IRocio RollanAún no hay calificaciones
- Resumen Biocel Solemne 2Documento25 páginasResumen Biocel Solemne 2Josefa CastilloAún no hay calificaciones
- Resumen Fisiologia Celular P1Documento15 páginasResumen Fisiologia Celular P1gallardo04Aún no hay calificaciones
- 6 - BiomembranasDocumento17 páginas6 - BiomembranasFlor BovettiAún no hay calificaciones
- Aparato de Golgi. Secreción y Endosomas NuevoDocumento53 páginasAparato de Golgi. Secreción y Endosomas NuevoFelipe MartinsAún no hay calificaciones
- Capitulo VI. Transporte Vesicular de Las Rutas SecretorasDocumento12 páginasCapitulo VI. Transporte Vesicular de Las Rutas SecretorasCamila Paulina Aravena SotoAún no hay calificaciones
- Biocel Med S1Documento6 páginasBiocel Med S1ferlagosmontecinosAún no hay calificaciones
- Estructura y Funciones de La Membrana CelularDocumento2 páginasEstructura y Funciones de La Membrana CelularFederica BirrielAún no hay calificaciones
- T2 - Membrana PlasmáticaDocumento13 páginasT2 - Membrana PlasmáticaEryka Ruiz AngueAún no hay calificaciones
- Resumen Segundo Parcial de IBMC 2Documento45 páginasResumen Segundo Parcial de IBMC 2belen100% (2)
- Proteinas Biologia LauDocumento14 páginasProteinas Biologia LauRita TolabaAún no hay calificaciones
- 1 de 2 CLASE Estructura y Funcion Mito, RER, Golgi, Liso ENBI-12 2023Documento9 páginas1 de 2 CLASE Estructura y Funcion Mito, RER, Golgi, Liso ENBI-12 2023Alanis Portillo RawlinsAún no hay calificaciones
- Unidad 2 Solemne Biologia Celular Rapaso - CompressDocumento40 páginasUnidad 2 Solemne Biologia Celular Rapaso - CompressabbysmilexpAún no hay calificaciones
- CUESTIONARIO TEMA 5Documento4 páginasCUESTIONARIO TEMA 5belenfmalvar73Aún no hay calificaciones
- Sistema de Membranas Citoplasmaticas o Sistema de EndomembranasDocumento9 páginasSistema de Membranas Citoplasmaticas o Sistema de EndomembranasValentina CelaniAún no hay calificaciones
- (AP) La Permeabilidad de Membrana PlasmáticaDocumento10 páginas(AP) La Permeabilidad de Membrana PlasmáticaNICOLE MARIAN GONZÁLEZAún no hay calificaciones
- Transporte Por VesículasDocumento3 páginasTransporte Por VesículasAnnaAún no hay calificaciones
- C11 - Sistemas de EndomembranasDocumento8 páginasC11 - Sistemas de EndomembranaskevinshustterAún no hay calificaciones
- C8 - Estructura y Función de Las Membranas CelularesDocumento7 páginasC8 - Estructura y Función de Las Membranas CelulareskevinshustterAún no hay calificaciones
- Tema 5 Tráfico VesicularDocumento44 páginasTema 5 Tráfico VesicularcarucastAún no hay calificaciones
- Fisiología CelularDocumento2 páginasFisiología CelularEduardo Jahir AnguloAún no hay calificaciones
- Componentes MolecularesDocumento25 páginasComponentes MolecularesYasbenia Araujo100% (1)
- Clase 10Documento22 páginasClase 10sebastiandiazgomesAún no hay calificaciones
- Evaluacion 13. Autoevaluacion Estructura y Funcion de La Membrana CelularDocumento5 páginasEvaluacion 13. Autoevaluacion Estructura y Funcion de La Membrana CelularAngely PaolaAún no hay calificaciones
- Sistema de EndomembranasDocumento13 páginasSistema de EndomembranasDiegoArenasCataldoAún no hay calificaciones
- Reticulo EndoplasmaticoDocumento20 páginasReticulo EndoplasmaticoDayanna Katherine Vasquez EscobarAún no hay calificaciones
- Certamen II Bio CelDocumento11 páginasCertamen II Bio CelAlejandra MartinezAún no hay calificaciones
- Orgánulos CelularesDocumento21 páginasOrgánulos Celulares30211202046Aún no hay calificaciones
- Organelos CelularesDocumento39 páginasOrganelos CelularesJorge TristanAún no hay calificaciones
- Tema 4.2Documento52 páginasTema 4.2Juancar Martinez CrespoAún no hay calificaciones
- Hidrolasas AcidasDocumento2 páginasHidrolasas AcidasLorena Mora Palomino100% (1)
- Teorico TP 10 1 BioDocumento7 páginasTeorico TP 10 1 BioCaterina Maurin 8 6 3 7 5 / 4Aún no hay calificaciones
- KMMA S9 Biologia CelularDocumento9 páginasKMMA S9 Biologia CelularEstefani Jissel Moreno LópezAún no hay calificaciones
- MEMBRANASDocumento32 páginasMEMBRANASHeydi JimenezAún no hay calificaciones
- TEMA 5 BiologíaDocumento8 páginasTEMA 5 BiologíamerxeabcAún no hay calificaciones
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012De EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Aún no hay calificaciones
- Lección 4. Célula Eucariótica (Grupo B)Documento7 páginasLección 4. Célula Eucariótica (Grupo B)Gabriel Shiner NoriegaAún no hay calificaciones
- Ciencias Zapandí Fascículo N 1Documento83 páginasCiencias Zapandí Fascículo N 1JosueCambronero50% (6)
- Nombre: Curso: Fecha:: Guía - Organelos CelularesDocumento8 páginasNombre: Curso: Fecha:: Guía - Organelos CelularesGuisellaAún no hay calificaciones
- Célula para ImprimirDocumento46 páginasCélula para ImprimirOrnella CarballoAún no hay calificaciones
- Atlas Sistema EndomembranaDocumento14 páginasAtlas Sistema EndomembranaLuis Diego Flores M.Aún no hay calificaciones
- Tráfico CelularDocumento46 páginasTráfico Celulardeaigl14Aún no hay calificaciones
- Fichas Celula Animal y VegetalDocumento3 páginasFichas Celula Animal y Vegetalariana alvarezAún no hay calificaciones
- Organelos CelularesDocumento19 páginasOrganelos CelularesmarcelaAún no hay calificaciones
- MICROVESICULASDocumento12 páginasMICROVESICULASWalter Bladimir Rivera JuarezAún no hay calificaciones
- FICHA 13 EXPLICA-citologíaDocumento8 páginasFICHA 13 EXPLICA-citologíaManuel Trujillo machacaAún no hay calificaciones
- Album Estructural de La CelulaDocumento8 páginasAlbum Estructural de La CelulaElmer ER0% (1)
- Biología CelularDocumento66 páginasBiología CelularMarco AguilarAún no hay calificaciones
- Actividades Tema II de Biología, LA CELULADocumento14 páginasActividades Tema II de Biología, LA CELULASaturnina MercedesAún no hay calificaciones
- T1-RESUMEN La Célula Como Unidad Estructural Relacionada A La Fisiología.Documento3 páginasT1-RESUMEN La Célula Como Unidad Estructural Relacionada A La Fisiología.Robin MalpartidaAún no hay calificaciones
- Celulas Eucariotas y ProcarotasDocumento57 páginasCelulas Eucariotas y ProcarotasFrank AdvAún no hay calificaciones
- Vías Por Las Que Llegan Las Moléculas A Los Lisosomas PDFDocumento1 páginaVías Por Las Que Llegan Las Moléculas A Los Lisosomas PDFDeliaM.CalderónAún no hay calificaciones
- Sistema Endomembranoso InfografíaDocumento1 páginaSistema Endomembranoso InfografíaLaura APAún no hay calificaciones
- Fundamentos Celulares Usamedic PrintDocumento16 páginasFundamentos Celulares Usamedic PrintjdiazpaAún no hay calificaciones
- BIO 5.2 Aparato de GolgiDocumento8 páginasBIO 5.2 Aparato de GolgiCarmen LostalAún no hay calificaciones
- Tema 3 LA CELULADocumento99 páginasTema 3 LA CELULAIVAN VLADIMIR ROCABADO ROCABADOAún no hay calificaciones
- Recop. Parciales BF HistoDocumento36 páginasRecop. Parciales BF HistoM PAún no hay calificaciones
- GLUCOSILACIONDocumento15 páginasGLUCOSILACIONGeraldine MendozaAún no hay calificaciones
- PDF-UNIDAD3 Ensomembranas 2018Documento54 páginasPDF-UNIDAD3 Ensomembranas 2018Gaby HerreraAún no hay calificaciones
- Unidad 4 - Actividad 2 Aparato de Golgi - José Alberto Fajardo Pérez - Ichabetel Adriana Tavares LunaDocumento16 páginasUnidad 4 - Actividad 2 Aparato de Golgi - José Alberto Fajardo Pérez - Ichabetel Adriana Tavares LunaElin MoralesAún no hay calificaciones
- Expo Ce LulaDocumento28 páginasExpo Ce Lulasara mosqueraAún no hay calificaciones
- Nivel CelularDocumento60 páginasNivel CelularJhisel100% (1)
- Introduccion A La Histologia.1Documento77 páginasIntroduccion A La Histologia.1Evelin Ramos100% (1)
- Taller 3 Celulas Animales y VegetalesDocumento16 páginasTaller 3 Celulas Animales y VegetalesRUT HELLENA BATISTA NARVAEZ ESTUDIANTE ACTIVOAún no hay calificaciones