
Ctenophora
| Ctenóforos | ||
|---|---|---|
| Rango temporal: Cámbrico-Reciente | ||
 Ctenóforo bioluminiscente y tentaculado no identificado. | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Subreino: | ¿Eumetazoa?, Ctenozoa | |
| Filo: |
Ctenophora Eschscholtz, 1829 | |
| Clases | ||


Los ctenóforos (Ctenophora, del griego Κτενός ctenos, «peine», y φόρος phoros, «portador») (pronunciación: [tenóforos]) son un filo de animales diblásticos[1] caracterizados por la presencia de unas células especializadas, los coloblastos, que producen una sustancia pegajosa utilizada para capturar las presas. Son exclusivamente marinos y se han descrito 166 especies.[2] Aunque poco conocidos, son muy abundantes en los mares de todo el mundo constituyendo una elevada proporción de la biomasa del plancton. Algunos tienen apariencia parecida a medusas, motivo por el cual fueron agrupados con ellas en el antiguo filo de los celentéreos, y otros parecen gusanos aplanados en los fondos oceánicos (formas reptantes). Viven desde la superficie hasta 3000 metros o más de profundidad.
Características generales
[editar]Muchos ctenóforos son similares a medusas, pero a diferencia de éstas que poseen cnidocitos, los ctenóforos poseen coloblastos, células exclusivas de este grupo, utilizadas para atrapar el alimento (zooplancton). Tienen simetría birradial o falsa simetría bilateral, presentando dos ejes de simetría. Unos pocos presentan simetría bilateral, pero no es único dentro de los radiados ya que en el filo Cnidaria también aparecen algunos casos con ese tipo de simetría.
Contienen ocho hileras lineales de cilios, denominados paletas natatorias o peines (de donde deriva su nombre) que se mueven sincrónicamente para nadar (los que lo hacen; algunos viven sobre el fondo o sobre otros animales). Algunos poseen un par de tentáculos, otros no. No tienen aparatos excretor, respiratorio ni circulatorio; sí poseen aparato digestivo y sistemas nervioso y muscular.
Tienen bioluminiscencia que depende de la mesoglea, donde además están las células musculares para las contracciones.
Su cuerpo, de aspecto gelatinoso, es esferoidal o piriforme; algunos tienen forma de cinta. En el polo oral se abre la boca, con forma de hendidura, y en el polo aboral (extremo opuesto) se encuentra un órgano sensorial característico que incluye un estatocisto.
Anatomía
[editar]Simetría
[editar]Los ctenóforos presentan generalmente una simetría que puede definirse como simetría birradial o bisimetría: dos planos dividen al animal en dos imágenes especulares a lo largo del eje oral-aboral. Boca y faringe están comprimidas, definiendo un plano de simetría bilateral al que llamamos plano bucal o sagital. La posición de los tentáculos define el segundo plano de simetría bilateral: plano tentacular o transversal, que es perpendicular al anterior.
Algunos autores consideran que en realidad los ctenóforos no tienen simetría radial ni bilateral, sino simetría rotacional, es decir, no existe un plano que los divida en imágenes especulares, como sí sucede en animales con simetría bilateral o radial; en su lugar, cualquier plano que se dibuje a través del eje oral-aboral central divide un ctenóforo en dos mitades iguales, pero simplemente giradas 180 grados.[3]
Aparato digestivo
[editar]A continuación de la boca hay una faringe que segrega enzimas y produce una pre digestión. A continuación aparece una zona ancha y amplia, el estómago, que se ramifica en 5 canales aumentando así su superficie para captar más agua con alimento. De los 5 canales, dos son los canales faríngeos, que parten del estómago regresando hacia la boca, pero antes de alcanzarla acaban en un fondo ciego. Otros dos canales son los canales transversales, de corta longitud, que a su vez se dividen en tres ramas, la primera es el canal tentacular (llamado así por acabar junto a las bolsas tentaculares) las otras dos ramas son los canales interradiales, que se vuelven a dividir en dos y acaban en los canales meridionales. Presentan ocho canales meridionales laterales, que discurren en dirección oral-aboral, donde se alojan las gónadas. Finalmente, a continuación del estómago, el canal aboral llega hasta la parte superior, al recto, y desemboca en dos poros anales que comunican el digestivo con el exterior.
Sistema nervioso
[editar]Es un plexo (entramado) subepidérmico de neuronas debajo de la dermis y está repartido por todo el cuerpo. Junto con el órgano sensorial apical, controla el inicio y fin de batido de las paletas natatorias, así como su ritmo, dirección del golpe eficaz y velocidad de onda.
Sistema locomotor
[editar]Se compone de ocho bandas meridianas de paletas natatorias, agrupadas en cuatro pares (cada par situado en un interradio), que parten del poro aboral y recorren el cuerpo hasta las inmediaciones del polo oral. Estas bandas poseen células ciliadas, cuyos cilios están parcialmente fusionados en forma de peine que se mueven batiendo juntos como una unidad. Estos peines son la característica que da nombre al filo; ctenos significa peine en griego. Las paletas se comunican en el polo aboral con el órgano del equilibrio o estatocisto. Cuatro surcos ciliados se prolongan en el exterior del estatocisto, bifurcándose para continuarse en las ocho bandas ciliadas.
El batido coordinado de las bandas ciliadas propulsa al animal por el agua. Las bandas de cilios de cada hilera baten en secuencia. Normalmente el golpe eficaz se da en sentido aboral, de forma que el desplazamiento sea con la boca por delante. Ante un estímulo lo bastante fuerte, todas las bandas pueden batir en sentido contrario (de modo que el animal nade en sentido contrario), o bien pueden batir en sentido contrario sólo unas hileras (para hacer girar el cuerpo).
Reproducción y desarrollo
[editar]Casi todos los ctenóforos son hermafroditas (monoicos), es decir, lo que en zoología se conoce por tener un mismo individuo los dos sexos y la capacidad de producir gametos femeninos y masculinos. Las gónadas producen gametos que se expulsan por el orificio bucal y la fecundación se produce en el agua. Da una larva cidipedia, plactónica ciliada y con tentáculos que originará un adulto.
La embriogenia recuerda en algunos aspectos a la de los protóstomos. La segmentación es total y determinada. La gastrulación tiene lugar por epibolia e invaginación. El mesénquima parece derivar del endodermo, como en los cnidarios, y por lo tanto debe considerarse como ectomesodermo. La gástrula se desarrolla pronto en una larva cidipedia, esférica u ovoidea, que nada libremente. Las especies planas de ctenóforos también poseen larvas de este tipo, lo que confirmaría que los ctenóforos ovoides son más primitivos.
Taxonomía
[editar]Hay más de 150 especies reconocidas aunque se estima que sea probablemente el filo animal con un número mayor de especies por descubrir debido a su vida planctónica en los fondos oceánicos, de muy difícil acceso. Tradicionalmente los ctenóforos se dividen en dos clases en función de la ausencia o presencia de tentáculos en los adultos:[4]
- Clase Tentaculata (o Tentaculifera) - tentaculados
- Contiene de lejos la mayor cantidad de especies conocidas; y como su nombre indica, poseen tentáculos, aunque a veces son vestigiales. Se dividen en ocho órdenes:
- Orden Cydippida, como Pleurobrachia pileus
- Orden Lobata
- Orden Platyctenida
- Orden Ganeshida
- Orden Cambojiida
- Orden Cryptolobiferida
- Orden Thalassocalycida
- Orden Cestida, como Cestum veneris
- Clase Nuda (o Atentaculata) - desnudos
- Orden Beroida - Se distinguen por la completa ausencia de tentáculos, como Beroe gracilis.
- Clase Scleroctenophora - Extintos del cámbrico que carecieron de tentáculos, tuvieron una coraza esclerotizada y ocho pares de filas de peine.[5]
Historia evolutiva
[editar]Los fósiles de ctenóforos, debido a su cuerpo blando y gelatinoso, son extraordinariamente raros, y los restos que se han interpretado como ctenóforos han sido hallados solo en lagerstätten, lugares en que el entorno estaba excepcionalmente condicionados para la preservación de tejidos blandos. Desde mitad de los años 1990 se han hallado solo dos especímenes en suficiente buen estado como para ser susceptibles de análisis, ambos del Devónico Inferior. Otras tres supuestas especies han sido halladas en el esquisto de Burgess y otras formaciones de similar antigüedad de Canadá, unos 505 millones de años (Cámbrico Medio); ninguna de ellas parece tener tentáculos pero tienen entre 24 y 80 hileras de peines, muchas más de las 8 hileras típicas de las especies actuales. Una de las especies posee una gran boca aparentemente rodeada por un repliegue que pudo haber sido muscular.[6] Evidencias provenientes de China sugieren que estos ctenóforos estaban ampliamente distribuidos en el Cámbrico, pero que eran muy distintos de los actuales, como por ejemplo una especie que tenía las hileras de peines sobre unas prominencias.[7]
Stromatoveris es un fósil del Cámbrico Inferior de Chengjiang (China), con una antigüedad de 525 millones de años; era sésil y tenía aspecto de fronde, y es muy similar a los vendobiontes del período precedente (Ediacárico). De-Gan Shu et al. hallaron en sus ramas lo que ellos consideran cilios, que el animal usaría para filtrar el alimento; sugieren que Stromatoveris es una "tía" evolutiva de los ctenóforos y que éstos se originaron a partir de formas sésiles similares, los descendientes de las cuales se hicieron nadadores y los cilios cambiaron su función alimenticia por una función propulsora.[8]
Se ha secuenciado el genoma de Mnemiopsis leidyi, un Bolinopsidae, en 2013, aportando claves de la evolución animal.[9]
En este cladograma se toma en cuenta la posible afinidad de trilobozoos, Eoandromeda, y los petalonamos con los ctenóforos, constituyendo el grupo denominado en este momento "Pan-Ctenophora" que engloba a los ctenóforos y a sus parientes extintos más cercanos que al resto de animales (esponjas (en sentido amplio), parahoxozoos y coanoflagelados (también en sentido amplio)), se había propuesto que si los ctenóforos de naturaleza pelágica fuesen descendientes de organismos bentónicos, se les podría considerar sucesores directos del aparentemente extinto filo Petalonamae, haciendo que dicho grupo no este realmente extinto.[10] Stromatoveris y Thaumaptilon representarían formas transicionales entre los petalonamos y los ctenóforos.
Se ha comprobado que los ctenóforos podrían ser el grupo hermano del resto de los animales, aunque se ha argumentado que podría ser por la atracción de ramas largas, pero la sintenía compartida de los animales rechaza fuertemente la hipótesis de la hermana porífera, en favor de la hermana ctenófora (véase Benthozoa),[11] no obstante los ctenóforos tampoco poseen genes para la formación de collares recubiertas de moco alrededor de flagelos (característica de los coanoflagelados, poríferos y parahoxozoos), por lo que podría ser incluso más basal que los coanoflagelados (por lo tanto divergir antes de la apomorfía de Choanoflagellata s.l), Gehlingia, un "trilobozoo" que se parece a los petalomanos en estructura puede ser también otro fósil transicional.[12]
Estudios sobre el sistema nervioso de los ctenóforos sugieren que podrían haber surgido independientemente,[13] y que estos aparecieron junto a los coloblastos.[14]
| Pan‑Ctenophora |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
Alternativamente se ha sugerido una afinidad de Trilobozoa con ParaHoxozoa, disolviendo definitivamente Vendobionta y Hexaradiozoa en favor de revivir a Coelenterata y Eumetazoa[15], dejando a los ctenóforos como los únicos descendientes de los petalonamos, implicando que la isomería es un rasgo convergente que apareció en Petalonamae, Trilobozoa y Proarticulata.[16]
Se recupera a Dinomischus, Daihua, y Xianguangia como un grupo monofilético denominado "Dinomischidae", siendo los dos últimos más cercanos entre sí que con Dinomischus, anteriormente considerado como un entoprocto de intestino en forma de U, ahora se le considera un ctenozoo con una cavidad digestiva ciega sin ano.[17][18], Siphusauctum se agrupa en su propia familia "Siphusauctidae" teniendo un aspecto globoso típico de los ctenóforos y posiblemente poros anales de 6 en lugar de solo 2 como los ctenóforos modernos.[17]
Un esqueleto está presente en los miembros bentónicos de ctenozoos, indicando la parafilia de Scleroctenophora.
| Pan‑Ctenophora |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||









.jpg)





Géneros extintos
[editar]Filogenia
[editar]En 1866 se propuso que los cnidarios y los ctenóforos estaban estrechamente relacionados porque ambos tenían una sola cavidad corporal que servía para las funciones digestivas, respiratorias y excretoras (cavidad gastrovascular), y se agruparon en el filo de los celenterados. En 1881 se propuso que los ctenóforos y los bilaterales guardaban una mayor relación ya que compartían características que no se dan en los cnidarios, como la presencia de musculatura en la capa central, mesoglea en ctenóforos y mesodermo en bilaterales. Análisis más recientes indican que dichas similitudes son bastante vagas, y el punto de vista actual, basado en datos moleculares, es que los cnidarios y los bilaterales están más relacionados entre sí que cada uno de ellos lo está con los ctenóforos. La agrupación formada por cnidarios y bilaterales se ha denominado Planulozoa, ya que sugiere que los primitivos bilaterales fueron similares a la larva plánula de los cnidarios.[19][20]
Dado que todos los ctenóforos actuales, excepto los beroidos, tienen una larva cidipedia, está ampliamente admitido que el último antecesor común a todos ellos era similar a los cidípedos, con una forma ovalada y un de tentáculos retráctiles. Los análisis basados en morfología concluyen que los cidípedos no son monofiléticos, con varias familias que resultan más cercanas a otros órdenes de ctenóforos.[21] Estudios más recientes basados en datos moleculares llega a conclusiones similares y sostienen que el último antepasado común de los ctenóforos actuales tenía aspecto de cidípedo y es relativamente reciente y que, tal vez sobrevivió a la extinción masiva del Cretácico-Terciario (hace 65,5 millones de años) mientras el resto de linajes se extinguieron.[22] En el mismo estudio, y tras incluir otros filos animales se llega a la conclusión que los cnidarios están más cercanamente emparentados con los bilaterales que los ctenóforos, aunque la certeza no es muy alta.
Cladogramas
[editar]Relación con otros animales
[editar]Los ctenóforos son un grupo importante para entender la evolución de los animales. Estudios recientes habían sugerido que los ctenóforos representarían las formas más basales del reino animal lo que dificultaría probar como se originaron los tejidos en los animales ya que los esponjas no poseen esta característica. Sin embargo se ha demostrado que las razones por la que los ctenóforos aparecen como el grupo más basal de los animales en análisis es porque tienen una desviación en la secuencia de nucleótidos y aminoácidos lo que produce su mala colocación. Estudios recientes han podido encontrar las siguientes relaciones para los ctenóforos intentado evitar las atracciones de ramas largas:[23][24][25]
| Animalia |
| ||||||||||||||||||||||||
Relación de los grupos de Ctenophora
[editar]Cladograma que muestra la filogenia del filo Ctenophora sobre la base de datos morfológicos y moleculares del ADN codificante para el ARNr-18s:[22]
| Ctenophora |
| ||||||||||||||||||||||||||||||||||||||||||
Por otro lado, el análisis multigénico, que además del ARNr 18S usa también el ARNr 5.8S, el 28S, ADN espaciadores y proteínas, da el siguiente resultado aproximado:[26]
| Ctenophora |
| ||||||||||||||||||||||||||||||||||||
Galería
[editar]-
 Ctenophora de Haeckel
Ctenophora de Haeckel -

-

-
 Coeloplana, un ctenóforo bentónico (Platyctenida)
Coeloplana, un ctenóforo bentónico (Platyctenida) -

-

-
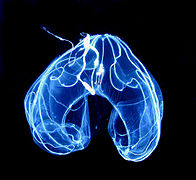
-







Ctenóforos extintos
[editar]-

-

-
 Archaeocydippida (Cydippida), del Devónico.
Archaeocydippida (Cydippida), del Devónico. -
 Ctenorhabdotus, del Cámbrico.
Ctenorhabdotus, del Cámbrico. -
 Stromatoveris, del Cámbrico.
Stromatoveris, del Cámbrico. -
 Sinoascus, del Cámbrico.
Sinoascus, del Cámbrico. -
 Fasciculus, del Cámbrico.
Fasciculus, del Cámbrico. -
 Xanioascus, del Cámbrico.
Xanioascus, del Cámbrico.



Referencias
[editar]- ↑ Brusca, R. C. & Brusca, G. J., 2005. Invertebrados, 2ª edición. McGraw-Hill-Interamericana, Madrid (etc.), XXVI+1005 pp. ISBN 0-87893-097-3.
- ↑ Chapman, A. D., 2009. Numbers of Living Species in Australia and the World, 2nd edition. Australian Biodiversity Information Services ISBN (online) 9780642568618
- ↑ Casey Dunn, S. Leys & S. Haddock 2015, The hidden biology of sponges and ctenophores. Trends in Ecology & Evolution. Volume 30, Issue 5, May 2015, Pages 282-291
- ↑ Mills, C.E. (15 de mayo de 2012). «Phylum Ctenophora: list of all valid scientific names». Consultado el 11 de diciembre de 2013.
- ↑ Qiang Ou et al. 2015, A vanished history of skeletonization in Cambrian comb jellies Science Advances 10 Jul 2015: Vol. 1, no. 6, e1500092 DOI: 10.1126/sciadv.1500092
- ↑ Conway Morris, S.; Collins, D. H. (marzo de 1996). «Middle Cambrian Ctenophores from the Stephen Formation, British Columbia, Canada». Philosophical Transactions of the Royal Society: Biological Sciences (Londres: The Royal Society) 351 (1337): 279-308. Consultado el 14 de febrero de 2009.
- ↑ Conway Morris, S. (2003). «The Cambrian “explosion” of metazoans and molecular biology: would Darwin be satisfied?». International Journal of Developmental Biology 47: 505-515. Consultado el 14 de febrero de 2009.
- ↑ Shu, D.-G.; Conway Morris, S., Han, J., Li, Y, et al. (2006). «Lower Cambrian Vendobionts from China and Early Diploblast Evolution». Science 312: 731-734. PMID 16675697. doi:10.1126/science.1124565. Consultado el 14 de febrero de 2009. (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- ↑ The Genome of the Ctenophore Mnemiopsis leidyi and Its Implications for Cell Type Evolution. Joseph F. Ryan, Kevin Pang, Christine E. Schnitzler, Anh-Dao Nguyen, R. Travis Moreland, David K. Simmons, Bernard J. Koch, Warren R. Francis, Paul Havlak, Stephen A. Smith, Nicholas H. Putnam, Steven H. D. Haddock, Casey W. Dunn, Tyra G. Wolfsberg, James C. Mullikin, Mark Q. Martindale, and Andreas D. Baxevanis. Science 13 December 2013: Vol. 342 no. 6164 DOI: 10.1126/science.1242592
- ↑ «Origen evolutivo de la asimetría en animales metazoarios primitivos». doi:10.1016/B978-008043404-9/50012-7.
- ↑ «Los enlaces de genes antiguos respaldan a los ctenóforos como hermanos de otros animales». Nature. 17 de mayo de 2023. doi:10.1038/s41586-023-05936-6.
- ↑ Paleontología dinámica: uso de la cuantificación y otras herramientas para descifrar la historia de la vida. Springer. 10 de junio de 2016. ISBN 9783319227771.
- ↑ «La red de nervios sincitiales en un ctenóforo agrega información sobre la evolución de los sistemas nerviosos». Science. 20 de abril de 2023. PMID 37079688. doi:10.1126/ciencia.ade5645. Consultado el 24 de abril de 2023.
- ↑ «Los tentáculos pegajosos de las medusas de peine pueden haberse revelado cuando las células nerviosas evolucionaron por primera vez». Science. 10 de enero de 2019. doi:10.1126/science.aaw6288.
- ↑ «Procesos de desarrollo de macrofósiles de Ediacaria». The Royal Society Publishing. 24 de febrero de 2021. ISSN 3362-2124. PMID 33622124. doi:10.1098/rspb.2020.3055.
- ↑ «Procesos de desarrollo de macrofósiles de Ediacaria». The Royal Society Publishing. 24 de febrero de 2021. ISSN 3362-2124. PMID 33622124. doi:10.1098/rspb.2020.3055.
- ↑ a b «Sésiles del Cámbrico, Ctenóforos del grupo de tallos que se alimentan en suspensión y evolución del plan corporal de gelatina en forma de peine». Current Biology. 21 de marzo de 2019. PMID 30905603. doi:10.1016/j.cub.2019.02.036.
- ↑ «Naturaleza tentacular de la 'columna' del diploblástico cámbrico Xiangangia sinica». Revista de Paleontología Sistemática. 27 de junio de 2022. ISSN 1477-2019. doi:10.1080/14772019.2023.2215787. Consultado el Aceptado el 11 de mayo de 2023, Publicado en línea 19 de junio de 2023.
- ↑ Wallberg, A., Thollesson, M., , Farris, J.S., and Jondelius, U. (2005). «The phylogenetic position of the comb jellies (Ctenophora) and the importance of taxonomic sampling». Cladistics 20: 558-578. doi:10.1111/j.1096-0031.2004.00041.x.
- ↑ Collins, A.G. (2002). «Phylogeny of Medusozoa and the Evolution of Cnidarian Life Cycles» (PDF). Journal of Evolutionary Biology 15 (3): 418-432. doi:10.1046/j.1420-9101.2002.00403.x. Archivado desde el original el 22 de septiembre de 2006. Consultado el 27 de noviembre de 2008.
- ↑ Harbison, G.R. (1985). «On the classification and evolution of the Ctenophora». En Conway Morris, S., George, J.D.,. Gibson, R., and Platt, H.M., ed. The Origins and Relationships of Lower Invertebrates. Clarendon Press. pp. 78-100. ISBN 019857181X.
- ↑ a b Podar, M.; Haddock, S.H.D, Sogin, M.L., and Harbison, G.R. (noviembre de 2001). «A Molecular Phylogenetic Framework for the Phylum Ctenophora Using 18S rRNA Genes». Molecular Phylogenetics and Evolution (Academic Press) 21 (2): 218-230. doi:10.1006/mpev.2001.1036. Archivado desde el original el 16 de mayo de 2008. Consultado el 14 de febrero de 2009.
- ↑ Feuda, Roberto; Dohrmann, Martin; Pett, Walker; Philippe, Hervé; Rota-Stabelli, Omar; Lartillot, Nicolas; Wörheide, Gert; Pisani, Davide (2017). «Improved Modeling of Compositional Heterogeneity Supports Sponges as Sister to All Other Animals». Current Biology (en inglés) 27 (24): 3864-3870.e4. PMID 29199080. doi:10.1016/j.cub.2017.11.008.
- ↑ Pisani, Davide; Pett, Walker; Dohrmann, Martin; Feuda, Roberto; Rota-Stabelli, Omar; Philippe, Hervé; Lartillot, Nicolas; Wörheide, Gert (15 de diciembre de 2015). «Genomic data do not support comb jellies as the sister group to all other animals». Proceedings of the National Academy of Sciences 112 (50): 15402-15407. Bibcode:2015PNAS..11215402P. PMC 4687580. PMID 26621703. doi:10.1073/pnas.1518127112.
- ↑ Kapli, Paschalia; Telford, Maximilian J. (11 Dec 2020). «Topology-dependent asymmetry in systematic errors affects phylogenetic placement of Ctenophora and Xenacoelomorpha». Science Advances 6 (10). doi:10.1126/sciadv.abc5162. Consultado el 17 de diciembre de 2020.
- ↑ Luis Arteaga et al. Revisiting the phylogeny of phylum Ctenophora: a molecular perspective F1000 Research 5:2881· También aquí
Enlaces externos
[editar] Wikimedia Commons alberga una galería multimedia sobre Ctenophora.
Wikimedia Commons alberga una galería multimedia sobre Ctenophora. Wikispecies tiene un artículo sobre Ctenophora.
Wikispecies tiene un artículo sobre Ctenophora.- Ctenophores from the São Sebastião Channel, Brazil Archivado el 4 de febrero de 2012 en Wayback Machine.
| Control de autoridades |
|
|---|
 Datos: Q102778
Datos: Q102778- Multimedia: Ctenophora / Q102778
- Especies: Ctenophora