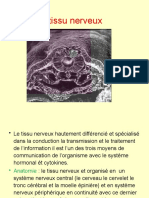
07-Le Tissu Nerveux
07-Le Tissu Nerveux
Télécharger au format pdf ou txt
Vous aimerez peut-être aussi
- Le Tissu NerveuxDocument10 pagesLe Tissu NerveuxANDRIANOME tafitaPas encore d'évaluation
- 08 - Le Tissu NerveuxDocument7 pages08 - Le Tissu NerveuxlahlahazouPas encore d'évaluation
- 4eme Type Tissu NerveuxDocument36 pages4eme Type Tissu Nerveuxgroupe_svi3Pas encore d'évaluation
- Résumé G.Chakib NerveuxDocument5 pagesRésumé G.Chakib NerveuxMaroua AbzPas encore d'évaluation
- Tissu NerveuxDocument6 pagesTissu Nerveuxtim SssPas encore d'évaluation
- Tissu Nerveux UCADDocument66 pagesTissu Nerveux UCADDiabel AndøPas encore d'évaluation
- Chapitre II: Organisation Cellulaire Du Système NerveuxDocument36 pagesChapitre II: Organisation Cellulaire Du Système Nerveuxnesrine MoadPas encore d'évaluation
- Leçon 03 Structure Du Tissu NerveuxDocument6 pagesLeçon 03 Structure Du Tissu NerveuxfrancisthomasdiemePas encore d'évaluation
- Le Tissu Nerveux 2022Document90 pagesLe Tissu Nerveux 2022babacar98.04Pas encore d'évaluation
- Tissu NerveuxDocument6 pagesTissu NerveuxOussama FendaliPas encore d'évaluation
- Neurologie RévisionsDocument8 pagesNeurologie Révisionstompierret0Pas encore d'évaluation
- Tissu Nerveux 2024Document50 pagesTissu Nerveux 2024dianeaminata2006Pas encore d'évaluation
- Physiologie Du NeuroneDocument21 pagesPhysiologie Du NeuronelbawainaPas encore d'évaluation
- Tissu Nerveux 2024Document5 pagesTissu Nerveux 2024Tamer HarkatPas encore d'évaluation
- Le Tissu NerveuxDocument16 pagesLe Tissu NerveuxZizou HamdiPas encore d'évaluation
- Cours SVT Tle D&C 2013-2014Document17 pagesCours SVT Tle D&C 2013-2014sandwidibernard76Pas encore d'évaluation
- 1 Er Cour PhysiologieDocument13 pages1 Er Cour Physiologiederdiri imènePas encore d'évaluation
- MI3 Neurosciences Histologie Du Systeme Nerveux LavabreDocument17 pagesMI3 Neurosciences Histologie Du Systeme Nerveux LavabreDjallal HassaniPas encore d'évaluation
- Chapitre 1Document33 pagesChapitre 1koulabaflorence5Pas encore d'évaluation
- Organisation Du Tissu NerveuxDocument1 pageOrganisation Du Tissu Nerveuxseckoumar365Pas encore d'évaluation
- Leçon 3. Structure Du Tissu Nerveux Vu Au MicroscopeDocument2 pagesLeçon 3. Structure Du Tissu Nerveux Vu Au Microscopemarieme.hkbPas encore d'évaluation
- 01 - Le Tissu NerveuxDocument13 pages01 - Le Tissu NerveuxNesrinePas encore d'évaluation
- Definition Tissu Nerveux PagineDocument19 pagesDefinition Tissu Nerveux PaginemiriamzatumuaPas encore d'évaluation
- Le Tissu NerveuxDocument51 pagesLe Tissu NerveuxYouness KhalfaouiPas encore d'évaluation
- Le Tissu Nerveux - Vue GénéraleDocument67 pagesLe Tissu Nerveux - Vue GénéraleSiaka YodaPas encore d'évaluation
- Neurophysiologie 2Document49 pagesNeurophysiologie 2123kiwiPas encore d'évaluation
- Les Tissus NerveuxDocument9 pagesLes Tissus NerveuxZoulkiffiPas encore d'évaluation
- Cours Neurohisto PCEM1 2006-2007Document50 pagesCours Neurohisto PCEM1 2006-2007Wiwi LauraPas encore d'évaluation
- Physio LogieDocument41 pagesPhysio Logiehaloempire25Pas encore d'évaluation
- Tissus Musculaires Et NerveuxDocument36 pagesTissus Musculaires Et NerveuxibovyclaraPas encore d'évaluation
- Fiche 1, Bio, EB10, ch4, Act 3, CoursDocument2 pagesFiche 1, Bio, EB10, ch4, Act 3, Coursabolech748Pas encore d'évaluation
- Neuro Physio LogieDocument24 pagesNeuro Physio Logiefabiancuberogonzalez3Pas encore d'évaluation
- TISSU NERVEUX - DR LEBOUABIDocument32 pagesTISSU NERVEUX - DR LEBOUABIIkramilla Karamilla100% (1)
- Generalites IDocument11 pagesGeneralites IzqwjjbngxnPas encore d'évaluation
- La Neurophysiologie Cours 2013Document17 pagesLa Neurophysiologie Cours 2013friends diary100% (1)
- Tissu Nerveux - PolycopiéDocument10 pagesTissu Nerveux - PolycopiéAmina DrkPas encore d'évaluation
- Physiologie Du Nerf 2022 Mede UM6Document163 pagesPhysiologie Du Nerf 2022 Mede UM6Girl Ytb100% (1)
- Copie de CM Différenciation NeuronaleDocument48 pagesCopie de CM Différenciation Neuronaleq5xj2j62z9Pas encore d'évaluation
- Tissu Nerveux Part1 MelDocument4 pagesTissu Nerveux Part1 MelmelPas encore d'évaluation
- Cours SN DR NaoualiDocument75 pagesCours SN DR NaoualiKobePas encore d'évaluation
- Neuroanatomie Complet JustineDocument123 pagesNeuroanatomie Complet JustineMaria OlteanPas encore d'évaluation
- MOD6 Le Syste Me Nerveux Central - La Moelle e Pinie Re PDFDocument3 pagesMOD6 Le Syste Me Nerveux Central - La Moelle e Pinie Re PDFmedPas encore d'évaluation
- Tissu NerveuxDocument5 pagesTissu Nerveuxt.ezzraimi.inesPas encore d'évaluation
- Cours Physiologie Animale 1-1Document150 pagesCours Physiologie Animale 1-1Mina OussarPas encore d'évaluation
- 8-Tissu NerveuxDocument99 pages8-Tissu NerveuxAy ManPas encore d'évaluation
- 5 - Physiologie Du NeuroneDocument8 pages5 - Physiologie Du Neuronemintchimechak2Pas encore d'évaluation
- Tissus NerveuxDocument19 pagesTissus NerveuxGuenzet Akila100% (1)
- Chap II Tissu NerveuxDocument43 pagesChap II Tissu NerveuxbadamassimarwanePas encore d'évaluation
- 01-Physilogie Du Tissu NerveuxDocument49 pages01-Physilogie Du Tissu NerveuxBook HunterPas encore d'évaluation
- BIOLE102 Neurobiologie: LégendeDocument117 pagesBIOLE102 Neurobiologie: LégendeDaniela RiosPas encore d'évaluation
- Chapitre 9 - Histologie Du Tissu NerveuxDocument7 pagesChapitre 9 - Histologie Du Tissu NerveuxLaurianePas encore d'évaluation
- Neuro AnatomieDocument58 pagesNeuro AnatomieDaniel DariusPas encore d'évaluation
- Tissu NerveuxDocument62 pagesTissu Nerveuxhejermtiraoui12Pas encore d'évaluation
- D 2 Physiochap 2 Partie 1Document8 pagesD 2 Physiochap 2 Partie 1Fathi DergaaPas encore d'évaluation
- Le Système NerveuxDocument8 pagesLe Système NerveuxazizPas encore d'évaluation
- Physiologie NerveuseDocument35 pagesPhysiologie NerveuseESGIS SmartPas encore d'évaluation
- Le Tissu NerveuxDocument2 pagesLe Tissu NerveuxslimdamakPas encore d'évaluation
- Psychophysiologie IDocument139 pagesPsychophysiologie Iالاسراء و المعراجPas encore d'évaluation
- Chapitre 1.le Tissu Nerveux. CMDocument74 pagesChapitre 1.le Tissu Nerveux. CMHazem AbdennebiPas encore d'évaluation
- Neurobiologie: Les Grands Articles d'UniversalisD'EverandNeurobiologie: Les Grands Articles d'UniversalisPas encore d'évaluation
- MECANISMES DABSORPTION ET MOUVEMENT DES IONS KaidDocument5 pagesMECANISMES DABSORPTION ET MOUVEMENT DES IONS KaidBemmoussat AsmaPas encore d'évaluation
- Les VitaminesDocument13 pagesLes VitaminesBemmoussat AsmaPas encore d'évaluation
- L'étude Agro-Pédologique D'un Sol: Une Sortie Le But de La SortieDocument3 pagesL'étude Agro-Pédologique D'un Sol: Une Sortie Le But de La SortieBemmoussat AsmaPas encore d'évaluation
- Vigne de Table PowerDocument24 pagesVigne de Table PowerBemmoussat AsmaPas encore d'évaluation
- Betterave FourragereDocument85 pagesBetterave FourragereBemmoussat AsmaPas encore d'évaluation
- Aubergine WordDocument5 pagesAubergine WordBemmoussat Asma100% (1)
- 06-Les Tissus MusculairesDocument9 pages06-Les Tissus MusculairesBemmoussat AsmaPas encore d'évaluation
- 2 - Le Tissu CartilagineuxDocument5 pages2 - Le Tissu CartilagineuxBemmoussat AsmaPas encore d'évaluation
- TD N°1 en Biologie Moléculaire Et Génie GénétiqueDocument4 pagesTD N°1 en Biologie Moléculaire Et Génie GénétiqueBemmoussat AsmaPas encore d'évaluation
- Glossaire GénétiqueDocument13 pagesGlossaire GénétiqueBemmoussat AsmaPas encore d'évaluation
- Généralités Sur La CelluleDocument24 pagesGénéralités Sur La CelluleOumarou KontaPas encore d'évaluation
- CM BioDocument13 pagesCM BioLaurine COTE COLISSONPas encore d'évaluation
- Cytologie 3Document3 pagesCytologie 3Idrissou FmsbPas encore d'évaluation
- Bio Cell 1Document25 pagesBio Cell 1Simo AjdirPas encore d'évaluation
- Guide SVT SecondeDocument57 pagesGuide SVT SecondeRoland Fortuné Deguenonvo100% (3)
- A CompleterDocument28 pagesA CompleterAliou FayePas encore d'évaluation
- Cellule AnimaleDocument5 pagesCellule AnimaleabdihamabPas encore d'évaluation
- Myosites À InclusionsDocument10 pagesMyosites À InclusionsAbdou RahmouniPas encore d'évaluation
- BACTERIOLOGIE (Cours Paramédicale 1)Document30 pagesBACTERIOLOGIE (Cours Paramédicale 1)Almenaouar Adel100% (2)
- (Sba-Medecine - Com) Cytologie en QCMDocument86 pages(Sba-Medecine - Com) Cytologie en QCMYaya TraoréPas encore d'évaluation
- Cytosquelette 1Document77 pagesCytosquelette 1Eduard SimaderPas encore d'évaluation
- Pathologie Non TumoraleDocument59 pagesPathologie Non TumoraleEmmanuelPas encore d'évaluation
- L3 BM Systématique Partie I PDFDocument18 pagesL3 BM Systématique Partie I PDFDjamila HEZILPas encore d'évaluation
- La Cellule VCDocument16 pagesLa Cellule VCfrancetombal14Pas encore d'évaluation
- Biologie Cellulaire Annales - 2013-2014 À 2020-2021Document190 pagesBiologie Cellulaire Annales - 2013-2014 À 2020-2021vrgab25251Pas encore d'évaluation
- 2010TOU30308Document207 pages2010TOU30308Imanouila SilverPas encore d'évaluation
- CytosqueletteDocument3 pagesCytosqueletteFares BENHENDAPas encore d'évaluation
- Cours No 03-La Cellule (Suite)Document8 pagesCours No 03-La Cellule (Suite)Feriel Feriel0% (1)
- 1 Généralites Sur La CelluleDocument40 pages1 Généralites Sur La CelluleOumarou Konta100% (1)
- 6 - Appareil de GolgiDocument9 pages6 - Appareil de GolgiiklsatarfaouiPas encore d'évaluation
- COURS Culture in Vitro L3 PVE - 2020Document90 pagesCOURS Culture in Vitro L3 PVE - 2020Kodji n'cho Guy StephanePas encore d'évaluation
- Organite Cellulaire: Définition, Rôles Et Exemples Organite Cellulaire: Définition, Rôles Et ExemplesDocument4 pagesOrganite Cellulaire: Définition, Rôles Et Exemples Organite Cellulaire: Définition, Rôles Et Exemplesranimboulaabi18Pas encore d'évaluation
- Cellules - Metabolisme Cellules - Photosynthese - Respiration - Seconde SVT 2023Document9 pagesCellules - Metabolisme Cellules - Photosynthese - Respiration - Seconde SVT 2023tototomato 17100% (1)
- RibosomesDocument9 pagesRibosomesBël RoumiPas encore d'évaluation
- Le Réticulum Endoplasmique - 2Document45 pagesLe Réticulum Endoplasmique - 2ilyes.jilouPas encore d'évaluation
- Planche L8Document2 pagesPlanche L8Ahmadou KANDJIPas encore d'évaluation
- 6 - La Glande Mammaire (Dr. Touil) V 2014-2015Document7 pages6 - La Glande Mammaire (Dr. Touil) V 2014-2015Pace MakerPas encore d'évaluation
- Bouhidel-Membrane CellulaireDocument126 pagesBouhidel-Membrane CellulaireChibi ChanPas encore d'évaluation
- Paras I To LogieDocument81 pagesParas I To LogiediamondishigamisenkuPas encore d'évaluation
- Extraction Du Jus de Betterave (1) - 2Document79 pagesExtraction Du Jus de Betterave (1) - 2Salma ZakritePas encore d'évaluation