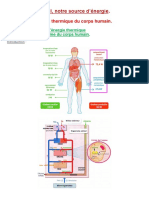
Bioénergétique & Biochimie Métabolique Chap 1
Bioénergétique & Biochimie Métabolique Chap 1
Télécharger au format pdf ou txt
Vous aimerez peut-être aussi
- Bioenergetique Et Principes de NutritionDocument174 pagesBioenergetique Et Principes de NutritionFarel DrbPas encore d'évaluation
- TSTL - Bioch.Cours .Chap.4.Metabolisme.I.Le - Metabolisme.energetiqueDocument8 pagesTSTL - Bioch.Cours .Chap.4.Metabolisme.I.Le - Metabolisme.energetiquekotobindenie0% (1)
- Photosynthèse: Les Grands Articles d'UniversalisD'EverandPhotosynthèse: Les Grands Articles d'UniversalisPas encore d'évaluation
- BioénergétiqueDocument6 pagesBioénergétiqueSalima ABDOUNIPas encore d'évaluation
- Biochimie MétaboliqueDocument7 pagesBiochimie MétaboliqueMBAÏADOUM NGARGUINAM RodriguePas encore d'évaluation
- Le Métabolisme ÉnergétiqueDocument5 pagesLe Métabolisme ÉnergétiqueJoses TatsiPas encore d'évaluation
- Cours 1 Biochimie MicrobienneDocument12 pagesCours 1 Biochimie Microbiennemo5049865Pas encore d'évaluation
- Biochimie1an Bioenergetique2022demmakDocument21 pagesBiochimie1an Bioenergetique2022demmakJunior MbatepPas encore d'évaluation
- 0.Polycopié Biochimie Mé TaboliqueDocument68 pages0.Polycopié Biochimie Mé Taboliquesaraqassid6Pas encore d'évaluation
- BioénergétiqueDocument16 pagesBioénergétiqueomarou18100% (6)
- Le Concept de La Bioénergétique 2020 2021-ConvertiDocument10 pagesLe Concept de La Bioénergétique 2020 2021-ConvertiJacques Ahishakiye100% (2)
- Bioénergétique S5 2025Document121 pagesBioénergétique S5 2025zoddanielndongPas encore d'évaluation
- Méta Cours 1 Et 2Document13 pagesMéta Cours 1 Et 2alicia.lrx76Pas encore d'évaluation
- Métabolisme Et BioénergétiqueDocument21 pagesMétabolisme Et BioénergétiqueRihab MaachouPas encore d'évaluation
- Bioenergetique 2021-22-1Document57 pagesBioenergetique 2021-22-1hanifaabou015Pas encore d'évaluation
- Cours de MétabolismeDocument22 pagesCours de MétabolismealmnaouarPas encore d'évaluation
- 8 - BioénergetiqueDocument12 pages8 - BioénergetiquePascal GadedjissoPas encore d'évaluation
- Bioénergétique - Cours 1Document6 pagesBioénergétique - Cours 1Youssra KambouaPas encore d'évaluation
- Biochimie Métabolique CHAP 1 Cours Métabolisme S4-2020-21Document30 pagesBiochimie Métabolique CHAP 1 Cours Métabolisme S4-2020-21khalid BOUBKRIPas encore d'évaluation
- BIO 201 - Cours 3Document25 pagesBIO 201 - Cours 3yroudwanePas encore d'évaluation
- Bioenerg Enzymo 2023Document91 pagesBioenerg Enzymo 2023Ghars Adam100% (1)
- Thermo 104656Document47 pagesThermo 104656Imene ImenePas encore d'évaluation
- BioénergetiqueDocument10 pagesBioénergetiqueFaculté De Médecine Béchar100% (7)
- BIOENERGETIQUE Sem 1Document5 pagesBIOENERGETIQUE Sem 1acnahinPas encore d'évaluation
- BioenergetiqueDocument50 pagesBioenergetiquemanelfor2004Pas encore d'évaluation
- Introduction Au MétabolismeDocument8 pagesIntroduction Au MétabolismeAtayiPas encore d'évaluation
- Biochimie MétaboliqDocument577 pagesBiochimie MétaboliqArcël RølPas encore d'évaluation
- Le Metabolisme Energetique DR GuedjatiDocument20 pagesLe Metabolisme Energetique DR GuedjatiMedMan689% (9)
- 2.bioener Metab RegulDocument140 pages2.bioener Metab RegulKenza Kassab100% (1)
- COURS 3 metabolismeDocument3 pagesCOURS 3 metabolismejulien.chandioux9Pas encore d'évaluation
- Biochimie Microbienne 1 1Document25 pagesBiochimie Microbienne 1 1khalilsassi90Pas encore d'évaluation
- BioE Des Rã©actions BiochimiquesDocument45 pagesBioE Des Rã©actions BiochimiquesSamuel G. SumoPas encore d'évaluation
- II-2-B) BilanDocument3 pagesII-2-B) BilanFELTENPas encore d'évaluation
- FC1 GALUSCA Rappels Des Grandes Voies Metaboliques Et Leurs Integrations 10.01Document20 pagesFC1 GALUSCA Rappels Des Grandes Voies Metaboliques Et Leurs Integrations 10.01Romane chapuisPas encore d'évaluation
- BC_3-1_Présentation_ThermodynamiqueDocument55 pagesBC_3-1_Présentation_Thermodynamiquekoukiee92Pas encore d'évaluation
- BIOENERGETIQUE CorrigéeDocument48 pagesBIOENERGETIQUE Corrigéesafemind100% (27)
- 1.introduction GeneraleDocument3 pages1.introduction Generaleadyam.messageriePas encore d'évaluation
- BIOENERGETIQUE Cours 2021-2022Document70 pagesBIOENERGETIQUE Cours 2021-2022Angel FahaPas encore d'évaluation
- Cours de Bioenergetique I - New1-MergedDocument469 pagesCours de Bioenergetique I - New1-MergedyvesrodrigezPas encore d'évaluation
- BioenergetiqueDocument29 pagesBioenergetiqueAlessandra LisiniPas encore d'évaluation
- 2.bioener Metab RegulDocument140 pages2.bioener Metab RegulKenza KassabPas encore d'évaluation
- II Alimentation Énergétique 2021Document21 pagesII Alimentation Énergétique 2021david davidPas encore d'évaluation
- Cours Respiration Fermentation Début regenerationATPDocument12 pagesCours Respiration Fermentation Début regenerationATPvalentinPas encore d'évaluation
- Cours - 1 Intoduction Pour La MétabolismeDocument6 pagesCours - 1 Intoduction Pour La Métabolismemiss anthropocenePas encore d'évaluation
- SV - S4 - Enz Et Metab - LaziriDocument52 pagesSV - S4 - Enz Et Metab - LaziriLinda KoundziPas encore d'évaluation
- Le Métabolisme ÉnergétiqueDocument33 pagesLe Métabolisme ÉnergétiquemimaPas encore d'évaluation
- BioénergétiqueDocument28 pagesBioénergétiqueshaiceark.bzhPas encore d'évaluation
- Bioénergétique Ndjamena 2022 2Document91 pagesBioénergétique Ndjamena 2022 2Mahamat Ali AbdramanPas encore d'évaluation
- cours Énergétique biologiqueDocument7 pagescours Énergétique biologiqueLONGTOUO JaurèsPas encore d'évaluation
- Synthèse Sur Les Réactions de Phospho-OxydationDocument33 pagesSynthèse Sur Les Réactions de Phospho-OxydationNoémie FrançoisPas encore d'évaluation
- Chap 4 Métabolisme FUE-SVT 2020-21.Ppt Partie 1Document56 pagesChap 4 Métabolisme FUE-SVT 2020-21.Ppt Partie 1Fatima zahrae AabidaPas encore d'évaluation
- Polycopie Voies Métaboliques S6Document42 pagesPolycopie Voies Métaboliques S6MOHSINA GHAMMATPas encore d'évaluation
- Introduction À La BioénergétiqueDocument108 pagesIntroduction À La Bioénergétiquebalypatricia33Pas encore d'évaluation
- cours-BIOCIMIE-METABOLIQUEpdfDocument5 pagescours-BIOCIMIE-METABOLIQUEpdffatmaezzhrarahmaniPas encore d'évaluation
- Métabolisme Et Bioénergétique 23-24-085953Document32 pagesMétabolisme Et Bioénergétique 23-24-085953LectorPas encore d'évaluation
- Les Deux Principes de La ThermodynamiqueDocument12 pagesLes Deux Principes de La ThermodynamiqueImene ImenePas encore d'évaluation
- 101-SN1-RE - Chapitre 4 - Théorie 2024Document26 pages101-SN1-RE - Chapitre 4 - Théorie 2024NioshaPas encore d'évaluation
- Sps Corps Humain 1 Charloux 08-02-21 35890Document16 pagesSps Corps Humain 1 Charloux 08-02-21 35890zt pas un fuguPas encore d'évaluation
- Medecine Bioenergetique PDFDocument19 pagesMedecine Bioenergetique PDFSoussou Souad100% (5)
- Études et recherches sur l'alimentation et le travail du cheval de troupeD'EverandÉtudes et recherches sur l'alimentation et le travail du cheval de troupePas encore d'évaluation
- Talus Et Son ÉquipeDocument42 pagesTalus Et Son ÉquipeAmjed LaritPas encore d'évaluation
- TD3 C3 Acides CarboxDocument15 pagesTD3 C3 Acides CarboxAbdou akim NdiayePas encore d'évaluation
- 8 Lois Newton Mouvement Circulaires UniformesDocument20 pages8 Lois Newton Mouvement Circulaires UniformesMilliardaire RüssëPas encore d'évaluation
- Pont RoulantDocument22 pagesPont RoulantAXIAL BIM INGINIERIEPas encore d'évaluation
- Cor RDM6Document7 pagesCor RDM6Chaker AmirPas encore d'évaluation
- Correction - TD SM2 Liaison CovalenteDocument11 pagesCorrection - TD SM2 Liaison CovalenteLo MomathPas encore d'évaluation
- Dépose-Repose de La Courroie de Distribution: o o o o o o o o o o o o o o o o o o o o o o o o oDocument5 pagesDépose-Repose de La Courroie de Distribution: o o o o o o o o o o o o o o o o o o o o o o o o oSEBPas encore d'évaluation
- 1 Année Master - Commande Electrique: ContinuousDocument2 pages1 Année Master - Commande Electrique: ContinuousMahmoud SettouPas encore d'évaluation
- Polycopie Travaux PratiqueDocument5 pagesPolycopie Travaux PratiqueAdil BahriPas encore d'évaluation
- Chap1 Etude Des Circuits Electriques en Regime Continu 1 1elt4Document88 pagesChap1 Etude Des Circuits Electriques en Regime Continu 1 1elt4angeloodjetychiquePas encore d'évaluation
- CC2Mec 1920Document2 pagesCC2Mec 1920elhaoudar.nassimaPas encore d'évaluation
- TP IPSecDocument6 pagesTP IPSecadem cherif100% (1)
- Statistique Chapitre2Document39 pagesStatistique Chapitre2Oussama Hammani100% (1)
- Exercice Circuit RLC SérieDocument3 pagesExercice Circuit RLC Sériealbymaiga18Pas encore d'évaluation
- Correction Série 2Document4 pagesCorrection Série 2ahmed chaouki Chami100% (4)
- Compte Rendu - Representation D'etatDocument20 pagesCompte Rendu - Representation D'etatabdelaziz.ouhtit-etuPas encore d'évaluation
- Cours06 Windows Server 2012 R2 Service DNSDocument29 pagesCours06 Windows Server 2012 R2 Service DNSSellé GueyePas encore d'évaluation
- Exercices Pompes HydrauliquesDocument9 pagesExercices Pompes HydrauliquesAnes BoudiaPas encore d'évaluation
- Compte Rendu Du TP2Document6 pagesCompte Rendu Du TP2Harabi AymenPas encore d'évaluation
- Simulation de La Fatigue Des Matériaux Par La Méthode Des Élément FinisDocument61 pagesSimulation de La Fatigue Des Matériaux Par La Méthode Des Élément FinisFrankie KOWAPas encore d'évaluation
- Corrigé Évaluation Comparaison - Demi-Droite Graduée - Vocabulaire de GéométrieDocument2 pagesCorrigé Évaluation Comparaison - Demi-Droite Graduée - Vocabulaire de Géométrieds.julia13Pas encore d'évaluation
- CoursDocument37 pagesCoursNicodeme SodeaPas encore d'évaluation
- Mémoire FINALEDocument106 pagesMémoire FINALECedrick KambPas encore d'évaluation
- Limites Cont TS1Document10 pagesLimites Cont TS1Roman BarbePas encore d'évaluation
- RankedDocument8 pagesRankedSali DouaePas encore d'évaluation
- TD 4: Algèbre Relationnelle: Bases de Données Modèle RelationnelDocument2 pagesTD 4: Algèbre Relationnelle: Bases de Données Modèle RelationnelMohamed amine AbdallahPas encore d'évaluation
- Electro 1ere Annee2002Document13 pagesElectro 1ere Annee2002Amadou SARRPas encore d'évaluation
- Fascicule RDMDocument21 pagesFascicule RDMFaustin Kouassi100% (1)
- FR GB Homologation Utac Feu Arriere x65 ph1 2 3 4Document14 pagesFR GB Homologation Utac Feu Arriere x65 ph1 2 3 4CristiSerseaPas encore d'évaluation
- Modélisation de La Ruine Fragile Et Ductile Des Poutres Renforcées Soumises À Un Chargement en FlexionDocument2 pagesModélisation de La Ruine Fragile Et Ductile Des Poutres Renforcées Soumises À Un Chargement en FlexionPatrick Saint-LouisPas encore d'évaluation