
Cnidaria

- une méduse Chrysaora melanaster ;
- une gorgone Annella mollis ;
- un corail Acropora cervicornis ;
- une anémone Nemanthus annamensis.
| Règne | Animalia |
|---|
Classes de rang inférieur
Les Cnidaires (Cnidaria) sont un embranchement d'animaux aquatiques (marins à 99 %, 1 % de dulcicoles seulement), possédant une symétrie radiale et des nématocystes (organites capables de lancer un harpon urticant pour attraper des proies). Cet embranchement regroupe notamment les anémones de mer, les méduses et les coraux.
L'appellation vient du grec ancien κνίδη / knídê, « ortie ») faisant allusion aux cellules urticantes caractéristiques de ces animaux (les cnidocytes ou cnidoblastes), le nom vernaculaire d'« orties de mer » étant donné par Aristote qui y regroupe les Acalèphes (méduses) et les Coralliaires[1]. Les zoologistes ont ainsi donné le nom de cet embranchement en hommage à Aristote[2].
Le terme de cœlentérés (Coelentera) désignait autrefois ce groupe mais inclut aussi l'embranchement voisin des cténaires. Les cnidaires sont bien représentés dans les fossiles : on les trouve jusque dans le Cambrien et peut-être même dès la faune de l'Édiacarien.
Les cnidaires existent sous deux formes : les formes fixées ou polypes (corail, anémone de mer) et les formes libres et mobiles (méduses). Il y a plus de 10 000 espèces reconnues.
Morphologie
[modifier | modifier le code]Aspect externe
[modifier | modifier le code]
Le corps des cnidaires se présente en un simple sac entourant une cavité gastrique qui s'ouvre vers l'extérieur par un pore unique qui a fonction de bouche et d'anus, entouré de tentacules (parfois régressés). Les formes polypes et méduses obéissent donc fondamentalement au même plan d'organisation, l'« ombrelle » des méduses correspondant au « pied » des formes fixées - il existe même des formes intermédiaires, comme les Staurozoa (méduses revenues à une vie fixe).
Le corps des cnidaires est organisé autour d'une symétrie radiale d'ordre pair (4 ou 6, ce qui détermine notamment le nombre des tentacules) : ils ressemblent donc à des « soleils » entourés de rayons. Chez certains anthozoaires, une symétrie biradiale s'ajoute secondairement à la symétrie radiaire[3]. Certains médusozoaires présentent une organisation bilatérale, conduisant certains chercheurs à suggérer que cette symétrie bilatérale trahit la condition ancestrale de l'embranchement des Cnidaria[4]. L'adulte est peu différent du stade embryonnaire.
Chez de nombreuses espèces (notamment le corail et de nombreux hydrozoaires), les polypes vivent en colonies (de polypes monomorphes ou polymorphes dont la division du travail se traduit par la présence de zoïdes différenciés et morphologiquement spécialisés)[5] qui rassemblent de très nombreux individus minuscules, connectés entre eux et pouvant sécréter un exosquelette calcaire très dur.
-
 Aurelia aurita, un cnidaire de forme méduse relativement rudimentaire.
Aurelia aurita, un cnidaire de forme méduse relativement rudimentaire. -
 L'anémone de mer Anthopleura elegantissima, un cnidaire de forme polype.
L'anémone de mer Anthopleura elegantissima, un cnidaire de forme polype. -
 Chez le corail (Acropora sp.), des polypes vivent en colonies et bâtissent un squelette calcaire commun.
Chez le corail (Acropora sp.), des polypes vivent en colonies et bâtissent un squelette calcaire commun. -
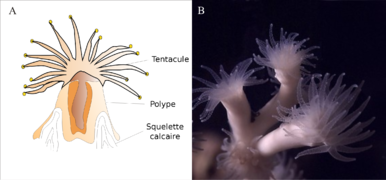 Polype de corail vu en coupe (Lophelia pertusa.
Polype de corail vu en coupe (Lophelia pertusa.




Anatomie
[modifier | modifier le code]
Les cnidaires sont des organismes diploblastiques c'est-à-dire qu'ils sont formés à partir de deux feuillets cellulaires embryonnaires seulement, l'endoderme et l'ectoderme (contrairement aux triploblastiques, qui en ont trois). Il peut y avoir entre ces deux feuillets une matrice, la mésoglée ou le mésenchyme selon le cas, qui ne constitue pas un véritable tissu cellulaire puisqu'il ne contient aucun organe différencié, mais où il existe un système nerveux relié aux cnidocytes (deux plexus nerveux, un sous-ectodermique et un sous-endodermique).
Le système nerveux, dérivé de l'ectoderme, est constitué d'un plexus, sans formation de ganglion ni de cerveau.
Le feuillet externe ou épiderme, d'origine ectodermique, est constitué de quatre sortes de cellules[6] :
- Les plus nombreuses sont les cellules myoépithéliales ectodermiques, disposées de façon longitudinale. Elles possèdent des fibres de muscles lisses en faisceau (myonèmes) de myosine qui permettent l'étirement et donc la rétractation. Ces cellules contiennent également des granules de sécrétion du mucus qui protège l'organisme.
- Les cellules sensorielles, qui, reliées à des fibres et cellules nerveuses, entraînent le déploiement du harpon des cellules urticantes (mouvement le plus rapide du règne animal).
- Les cellules interstitielles, isolées ou en amas. Généralement situées à la base des cellules myoépithéliales, elles restent à l'état de pluripotence et servent à remplacer des cellules mortes ou abîmées et sont aussi à l'origine des cellules germinales ou gonocytes.
- Les cnidocytes, cellules urticantes en forme de crochets. Elles contiennent une vésicule dérivée de l'appareil de Golgi remplie de poison. Le contenu de cette vésicule (actinogestine) est expulsé vers l'extérieur avec un harpon urticant lorsque le cnidocil (une expansion sensorielle excitable de la cellule) est touché. La cellule meurt après cette action.
- Éventuellement les cellules nerveuses, bien qu'elles ne soient pas inféodées à un tissu[réf. nécessaire].
Le feuillet interne ou endoderme, d'origine endodermique, aussi appelé gastroderme (lieu de la digestion) joue un rôle digestif ; il se compose des 4 types de cellules suivants[6] :
- Cellules myoépithéliales contenant des myonèmes qui ont une action perpendiculaire à celle des cellules myoépithéliales ectodermiques et aussi des vacuoles de pinocytose (digestion des éléments liquides, s'opposant à la phagocytose des éléments solides) et ayant des flagelles (afin d'amener la nourriture jusqu'à l'organisme);
- Cellules glandulaires contenant des granulations d'enzymes protéolytiques (destruction des protéines);
- Cellules sensorielles qui possèdent un cil excitable apical et des prolongements basaux qui possèdent des liaisons avec les neurones et les cellules myoépithéliales ;
- Cellules interstitielles
- Cellule basale destinée à remplacer les cellules de l'endoderme
Entre les deux couches principales, se trouve une couche intermédiaire de gelée anhiste, la mésoglée. Elle se compose principalement d'eau, mais il y existe des cellules nerveuses qui ont un rôle de coordination[7],[8]. La mésoglée est très importante chez les méduses et un peu moins chez les polypes.
Reproduction
[modifier | modifier le code]
En général, les cnidaires peuvent alterner entre forme polype et forme libre au cours de leur cycle reproductif, sauf les anthozoaires qui n'existent que sous la forme fixée (la larve est cependant planctonique). Le mode reproductif varie entre groupes, depuis l'alternance stricte polype/méduse à chaque génération jusqu'à la reproduction dans un seul des deux modes. Cependant pour un groupe donné une forme domine souvent nettement l'autre.
Les spermatozoïdes du mâle sont libérés dans son estomac avant d’être éjectés dans les eaux maritimes. La femelle garde ses ovules dans son estomac. La femelle ingurgite les spermatozoïdes portés par les courants marins et la fécondation va avoir lieu à l’intérieur même de l’estomac de la femelle. Les œufs ainsi créés donneront naissance à des larves appelées planula et qui correspondent au stade morula d'autres animaux plus complexes. La planula est entièrement recouverte de cils qui l’aident à se propulser jusque sur les lobes de la bouche de sa mère. C'est accrochées à ces lobes, par là où la nourriture passe, que les planulas vont grandir jusqu’à pouvoir nager toutes seules en pleine mer. Une fois qu’elle quitte la bouche de la mère, la planula va se fixer à un rocher ou bien à une algue. C’est là que la planula se transforme en polype avec des tentacules. Elle grandit ainsi jusqu’à ce que des sillons apparaissent autour de son corps. Ces sillons se creusent et le polype en vient à ressembler à une pile de saladiers. Le bloc du dessus bourgeonne avant de se détacher et de se transformer en larve appelée éphyra. C’est cette éphyra qui au fil du temps va devenir une méduse adulte.
Après gastrulation, cette larve se fixe et forme un polype. Le polype possède alors deux choix de reproduction : par clonage ou par émission de gamètes. Pour se cloner, il produit un stolon qui va servir de point de germination pour un nouveau polype. Les deux polypes restent liés par le stolon qui permet des échanges métaboliques. Ainsi se forme une colonie, sorte de super-organisme où les différents individus peuvent se spécialiser.
Lorsque les conditions (taille de la colonie, facteurs environnementaux) sont favorables, certains polypes se métamorphosent en méduses qui vont mener une vie pélagique, contrairement au polype qui est benthique. Les méduses peuvent se reproduire de façon végétative pour donner d'autres méduses, ou par des gamètes pour recommencer un nouveau cycle au stade polype.
Chaque polype peut se transformer en 6 à 8 méduses.
Ces deux formes sont liées par alternance de multiplication asexuée et reproduction sexuée qu'on appelle la métagenèse.
Nutrition
[modifier | modifier le code]
Opercule
Digitation réversible
/ / / Barbilles
Venin
Épiderme de la victime
Tissus de la victime.
La quasi-totalité des cnidaires sont carnivores, même si certains se complémentent par photosynthèse. Ils se nourrissent de proies venant au contact des tentacules, incluant du plancton, des protistes, divers vers, des crabes, d’autres cnidaires et même des poissons. Ils capturent et immobilisent les proies grâce à leurs tentacules recouvertes de cellules urticantes, les « cnidoblastes »[10] (cellules spécialisées comportant un appareil venimeux muni d’une sorte de harpon), à usage défensif ou de prédation[11]. Ces cellules peuvent produire des toxines anesthésiantes appelées actinogestines ou actino-congestines (polypeptides de 14 acides aminés), qui paralysent la proie harponnée[12]. Les tentacules apportent ensuite la proie vers la bouche. La digestion est d’abord extracellulaire : des cellules spécialisées sécrètent du mucus et des enzymes digestives dégradant la nourriture (un certain nombre de bactéries interviennent également dans le processus). Les particules alimentaires partiellement digérées sont ensuite pinocytées et la digestion se termine au niveau intracellulaire. Les résidus de la digestion sont évacués par la bouche qui sert également d'anus.
Écologie
[modifier | modifier le code]
Presque tous les cnidaires vivent en milieu marin mais les hydres sont rencontrées dans les eaux marines et dans les eaux douces (espèce parfois retrouvée en aquarium d'eau douce) selon les espèces.
Les cnidaires vivent très souvent en association symbiotique. Il s’agit souvent d’une endosymbiose avec des dinoflagellés du genre Symbiodinium appelés zooxanthelles. En absorbant le CO2 produit par le cnidaire, les algues utilisent l’énergie solaire par photosynthèse pour produire des glucides que le cnidaire utilise comme source de nutriments. Les zooxanthelles apportent une grande quantité d’énergie aux cnidaires. Elles favorisent par exemple la précipitation du carbonate de calcium et l’élaboration du squelette constituant les récifs coralliens. En contrepartie, le cnidaire offre une protection à son endosymbiote.
La relation entre l’anémone de mer et le poisson clown est un exemple d'association de type mutualisme. Un mucus protecteur sur le corps du poisson lui permet de tolérer le venin produit par l’anémone. Le poisson trouve un abri au sein de l’anémone. En contrepartie le poisson clown peut servir de leurre pour attirer des proies vers l’anémone. Il peut aussi défendre son anémone contre des attaques de certains prédateurs pouvant brouter l’anémone.
Classification
[modifier | modifier le code]Principaux groupes
[modifier | modifier le code]


.jpg)
Voici quelques caractéristiques des groupes principaux :
- Classe des Anthozoaires : représentés seulement par la forme polype, solitaires (anémone de mer) ou coloniaux (corail).
- sous-embranchement Medusozoa : comprend toujours un stade méduse
- Classe des Hydrozoaires : les Hydraires sont fixées sous la forme polype (Hydres) ou libres et solitaires sous la forme méduse.
- Classe des Scyphozoaires : polypes à l'état larvaire devenant des méduses à l'état adulte.
- Classe des Cubozoaires : méduses à forme plus ou moins cubique. Elles ont la particularité de posséder des yeux complexes.
- Classe des Staurozoaires : restent à l'état méduse mais fixé au substrat pendant toute leur vie.
- sous-embranchement des Myxozoaires : des parasites.
|
Selon World Register of Marine Species (5 février 2018)[13] :
|
Selon ITIS (20 février 2014)[14] :
|
Auxquels il faut ajouter des classes disparues comme :
- classe Cyclozoa †
-
 Arachnanthus nocturnus (cérianthe Penicilaria)
Arachnanthus nocturnus (cérianthe Penicilaria) -
 Cerianthus filiformis (cérianthe Spirularia)
Cerianthus filiformis (cérianthe Spirularia) -

-
 Cirrhipathes sp. (Antipatharia)
Cirrhipathes sp. (Antipatharia) -

-
 Acropora muricata (corail Scleractinia)
Acropora muricata (corail Scleractinia) -

-

-

-

-
 Carybdea branchi (cuboméduse Carybdeida)
Carybdea branchi (cuboméduse Carybdeida) -
 Chiropsalmus quadrigatus (cuboméduse Chirodropida)
Chiropsalmus quadrigatus (cuboméduse Chirodropida) -

-

-

-
-

-

-
 Crossota sp. (Trachymedusae)
Crossota sp. (Trachymedusae) -
-

-
 Atolla wyvillei (méduse Coronatae)
Atolla wyvillei (méduse Coronatae) -
 Cephea cephea (méduse Rhizostomeae)
Cephea cephea (méduse Rhizostomeae) -
 Aurelia aurita (méduse Semaeostomeae)
Aurelia aurita (méduse Semaeostomeae) -





.jpg)

_1.jpg)





_Blue_button_at_Thotlakonda_Beach_06.JPG)








.jpg)

_Luc_Viatour.jpg)

Phylogénie
[modifier | modifier le code]
La phylogénie des cnidaires reste un sujet de recherche ouvert. Marques & Collins (2004)[15], proposent pour les médusozoaires la phylogénie suivante :
N.B. : Les clades de parasites Myxozoa et Polypodiozoa n'ont pas été intégrés à cette étude.

Évolution
[modifier | modifier le code]Registre fossile
[modifier | modifier le code]Les premiers fossiles d'animaux largement acceptés sont des cnidaires d'apparence plutôt moderne, datant peut-être d'environ 580 millions d'années, bien que les fossiles de la formation de Doushantuo ne puissent être datés qu'approximativement. L'identification de certains d'entre eux comme des embryons d'animaux a été contestée, mais d'autres fossiles de ces roches ressemblent fortement à des tubes et à d'autres structures minéralisées faites par des coraux. Leur présence implique que les lignées cnidaires et bilatérales avaient déjà divergé. Bien que le fossile édiacarien Charnia ait été classé comme une méduse ou une plume de mer, une étude plus récente des modèles de croissance de Charnia et des cnidaires modernes a mis en doute cette hypothèse, laissant le polype canadien Haootia (en) et l'Auroralumina britannique comme les seuls fossiles de corps cnidaires reconnus de l'Édiacarien. Peu de fossiles de cnidaires sans squelette minéralisé sont connus à partir de roches plus récentes, sauf dans les lagerstätten qui ont conservé des animaux au corps mou.
Quelques fossiles minéralisés ressemblant à des coraux ont été trouvés dans des roches de la période cambrienne et des coraux se sont diversifiés au début de l'Ordovicien. Ces coraux, qui ont été anéantis lors de l'extinction Permien-Trias il y a environ 252 millions d'années, n'ont pas dominé la construction des récifs puisque les éponges et les algues ont joué un rôle prédominant. Au cours de l'ère mésozoïque, les bivalves rudistes étaient les principaux constructeurs de récifs, mais ils ont été anéantis lors de l'extinction Crétacé-Paléogène il y a 66 millions d'années, et depuis lors, les principaux constructeurs de récifs sont les coraux scléractiniens.
Place des cnidaires dans le règne animal
[modifier | modifier le code]- Les types d'organisation présentés ici sont des grades évolutifs ne correspondant généralement pas à des groupes monophylétiques, mais paraphylétiques (ne comportant pas tous les descendants d'un même ancêtre — exemple : les descendants d'ancêtres vermiformes ne sont pas tous aujourd'hui des vers, etc.).
- En jaune : les principales explosions radiatives.
| > Unicellulaires procaryotes (cellule sans noyau) | ♦ Échinodermes : Oursins, Crinoïdes, Concombres de mer, étoiles de mer et ophiures | ♦ Bivalves (coquillages) | ||||
| > Unicellulaires Eucaryotes (cellules à noyau) | ♦ Gastéropodes (escargots, limaces, etc.) | |||||
| > Éponges (organisme multicellulaire) | ♦ Mollusques | ♦ Céphalopodes (pieuvres, seiches) | ||||
| > Polype : hydres, coraux et méduses | ||||||
| > Vers (mobilité et tube digestif) bilatériens | ♦ Trilobites (de deux à 24 pattes — éteint) | |||||
| > Poissons agnathes (sans mâchoire) | ♦ Arthropodes primitifs type myriapodes (beaucoup de pattes) | ♦ Décapodes : crabes et écrevisses (dix pattes) | ||||
| > Poissons primitifs (poissons cartilagineux) | ♦ Arachnides : araignées, scorpions et acariens (huit pattes) | ♦ Libellules | ||||
| > Poissons typiques (poissons osseux) | ♦ Serpents | > Hexapodes (à six pattes) : Insectes type Apterygota (primitifs sans ailes) | ♦ Blattes, mantes, termites | |||
| > Poissons type Sarcopterygii (à nageoires charnues) | ♦ Dinosaures (éteint) | ♦ Orthoptères (sauterelles, grillons) | ||||
| > Tétrapodes primitifs (type Amphibiens) | ♦ Crocodiles | ♦ Marsupiaux | ♦ Hémiptères (punaises, cigales…) | |||
| > Reptiles primitifs (Amniotes de type Lézards) | ♦ Tortues | ♦ Insectivores (Taupes, Hérissons…) | ♦ Coléoptères (hannetons, coccinelles…) | |||
| ♦ Oiseaux | ♦ Chiroptères (Chauves-Souris) | ♦ Hyménoptères (abeilles, guêpes, fourmis) | ||||
| ♦ Primates | ♦ Diptères (mouches) | |||||
| > Mammifères primitifs type monotrèmes | ♦ Rongeurs et Lagomorphes (lapins) | ♦ Lépidoptères (papillons) | ||||
| ♦ Carnivores | ♦ | |||||
| ♦ Ongulés | ♦ |
Par rapport aux éponges, le passage à une organisation de type anémone répond à la question récurrente : comment se nourrir ? La formule mise au point par ce groupe consiste à pousser la nourriture vers un « ventre » (cavité gastrique) où elle est digérée sans pouvoir en sortir. Cette stratégie novatrice permet de se nourrir de proies plus grosses (que les éponges ne peuvent pas filtrer). Dans l'acquisition progressive de fonctionnalités animales, cette évolution suppose deux choses : les cellules se spécialisent (avec l'acquisition de cellules nerveuses et musculaires permettant des mouvements coordonnés) et l'organisme gagne la capacité à prendre une forme définie (morphogénèse), pour que des tentacules efficaces puissent pousser leur proie vers une cavité gastrique fonctionnelle.
Parmi les autres adaptations importantes qu'ont développées les cnidaires par rapport aux éponges, nous trouvons l'organisation cellulaire. En effet, les cnidaires furent les premiers organismes à posséder une réelle structure pluricellulaire. Contrairement aux éponges (qui constituent un ensemble de cellules seulement juxtaposées, possédant des fonctions différenciées[réf. nécessaire]), les cnidaires sont des organismes pluricellulaires (ensemble de cellules liées entre elles, appartenant au même organisme, possédant des fonctions différenciées et participant au métabolisme de l'individu). Les cnidaires ont donc inventé via cet accrochage intercellulaire les premiers tissus. Les cnidaires ont dû mettre au point par la même occasion un système permettant d'envoyer un message aux différentes cellules de l'organisme afin de coordonner leur action. Nous voyons ainsi apparaître les premières ébauches de systèmes nerveux et musculaire. On parle plutôt de « protocellules nerveuses » et de cellules myoépithéliales.
Les coraux représentent un stade intermédiaire, où les formes benthiques n'ont que des fonctionnalités de type spongiaire alors que les formes pélagiques adoptent une organisation fonctionnelle de type méduse. Au fil du temps, l'avantage évolutif de la réactivité sur la rigidité se fait sentir et l'organisation fonctionnelle des anémones émerge comme solution gagnante, ouvrant la voie aux bilatériens initialement vermiformes.
Phylogénie
[modifier | modifier le code]Il est difficile de reconstituer les premiers stades de l'arbre phylogénétique des animaux en se fiant uniquement la morphologie (leurs forme et structure), car les différences entre Éponges, Cnidaires et Cténophores, Placozoa et Bilateria (les animaux les plus complexes) sont telles qu'elles ne permettent guère de comparaisons. Par conséquent, la construction de l'arbre s'appuie désormais principalement sur la biologie moléculaire, qui regroupe les organismes en fonction des similitudes ou différences du point de vue biochimique, généralement dans leur ADN ou leur ARN[16].

On pense maintenant généralement que les Calcarea (éponges avec des spicules de carbonate de calcium) sont plus étroitement liées aux Cnidaria, Ctenophora et Bilateria (tous les animaux les plus complexes) qu'aux autres groupes d'éponges[17],[18],[19]. En 1866, il a été proposé que Cnidaria et Ctenophora étaient plus étroitement liés les uns aux autres qu'à Bilateria et formaient un groupe appelé Coelenterata (intestin en forme de cavité), car Cnidaria et Ctenophora dépendent tous deux du flux d'eau entrant et sortant d'une seule cavité, pour l'alimentation, l'excrétion et la respiration. En 1881, il a été proposé que Ctenophora et Bilateria étaient plus étroitement liés l'un à l'autre, car ils partageaient des caractéristiques qui manquent à Cnidaria, par exemple les muscles de la couche intermédiaire (mésoglée chez Ctenophora, mésoderme chez Bilateria). Cependant, des analyses plus récentes indiquent que ces similitudes sont plutôt vagues, et l'opinion actuelle, basée sur la phylogénétique moléculaire, est que Cnidaria est plus proche de Bilateria que de Ctenophora. Ce groupement de Cnidaria et de Bilateria a été nommé Planulozoa car il suggère que les premiers Bilatériens étaient apparentables aux larves dites planula des Cnidaires[20],[21].
Au sein des Cnidaires, les Anthozoa (anémones de mer et coraux) sont considérés comme le groupe frère du reste, ce qui suggère que les premiers cnidaires étaient des polypes sessiles sans stade de méduse. Cependant, on ne sait pas comment les autres groupes ont acquis le stade de la méduse, puisque les Hydrozoa forment des méduses en bourgeonnant du côté du polype tandis que les autres Medusozoa le font en les séparant de la pointe du polype. Les similitudes dans les doubles parois corporelles des Staurozoa et Conulariida (en) éteints suggèrent qu'ils sont étroitement apparentés[20],[22].
En 2005, Katja Seipel et Volker Schmid ont suggéré que cnidaires et cténophores seraient des descendants simplifiés d'animaux triploblastiques, puisque les cténophores et le stade méduse de certains cnidaires ont un muscle strié, qui chez les bilatériens provient du mésoderme[23].
Dans les analyses de phylogénie moléculaire à partir de 2005, d'importants groupes de gènes du développement montrent la même variété chez les cnidaires que chez les chordés[24]. En effet,[pas clair] les cnidaires, et surtout les anthozoaires (anémones de mer et coraux), conservent certains gènes présents chez les bactéries, les protistes, les plantes et les champignons mais pas chez les bilatériens[25].
Le génome mitochondrial des cnidaires médusozoaires, contrairement à ceux des autres animaux, est linéaire avec des gènes fragmentés[26]. La raison de cette différence est inconnue.
Voir aussi
[modifier | modifier le code]Références taxinomiques
[modifier | modifier le code]- (en) Référence WoRMS : Cnidaria Verrill, 1865 (+ liste classes + liste ordres)
- (en) Référence Paleobiology Database : Cnidaria Hatschek 1888
- (fr + en) Référence ITIS : Cnidaria Hatschek, 1888
- (en) Référence Animal Diversity Web : Cnidaria
- (en) Référence Tree of Life Web Project : Cnidaria
- (en) Référence Fauna Europaea : Cnidaria (consulté le )
Bibliographie
[modifier | modifier le code]- Grasse P. P., Doumenc D. Zoologie : 1. Invertébrés. Masson 3e éd. 1990
- (en) Stefano Goffredo et Zvy Dubinsky, The Cnidaria, past, present and future. The world of Medusa and her sisters, Springer, , 855 p. (ISBN 978-3-319-31303-0, lire en ligne)
- (en) Daly, M., Brugler, M. R., Cartwright, Paulyn, Collins, Allen Gilbert, Dawson, M. N., Fautin, D. G., France, S. C., McFadden, C. S., Opresko, Dennis M., Rodriguez, E., Roman, S. L., and Stake, J. L., « The phylum Cnidaria: A review of phylogenetic patterns and diversity 300 years after Linnaeus », Zootaxa, no 1628, , p. 127–182 (DOI 10.5281/zenodo.180149, lire en ligne)
Articles connexes
[modifier | modifier le code]- Cnidaria (classification phylogénétique)
- Photo-guide taxinomique du monde animal
- Photo-guide taxinomique de la faune et de la flore sous-marine
- Le lac aux méduses des Palaos
- Phytozoaire
Liens externes
[modifier | modifier le code]- Ressources relatives au vivant :
- Animal Diversity Web
- Australian Faunal Directory
- Dyntaxa
- EPPO Global Database
- EU-nomen
- Fauna Europaea
- Paleobiology Database
- Global Biodiversity Information Facility
- iNaturalist
- Interim Register of Marine and Nonmarine Genera
- Nederlands Soortenregister
- New Zealand Organisms Register
- Système d'information taxonomique intégré
- World Register of Marine Species
- Ressource relative à la santé :
- Notices dans des dictionnaires ou encyclopédies généralistes :
- Les Hydraires à La Réunion et dans l'océan Indien
- Les Cnidaires
- Biologie et développement des Cnidaires (Mer et littoral)
- Cours en ligne sur les cnidaires de l'UCLouvain
Notes et références
[modifier | modifier le code]- ↑ Jacqueline Goy et Anne Toulemont, Méduses, Musée océanographique, , p. 10
- ↑ Jacqueline Goy, « Les paradoxes des méduses », Pour la Science, no 299, , p. 37
- ↑ « Symétrie biradiale »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?), sur simulium.bio.uottawa.ca.
- ↑ (en) D.Q. Matus et al., « Molecular evidence for deep evolutionary roots of bilaterality in animal development », Proceedings of the National Academy of Sciences of the United States of America, vol. 103, no 30, , p. 11195–11200 (DOI 10.1073/pnas.0601257103).
- ↑ (en) P.S.Verma, Invertibrate Zoology, S. Chand Publishing, , p. 312.
- Daniel Richard, Patrick Chevalet, Sylvie Fournel, Nathalie Giraud, Frédéric Gros, Patrick Laurenti, Fabienne Pradere, Thierry Soubaya, Biologie, Dunod, , p. 95.
- ↑ Marijana Miljkovic-Licina, Dominique Gauchat et Brigitte Galliot, « Neuronal evolution: analysis of regulatory genes in a first-evolved nervous system, the hydra nervous system », Elsevier BV, vol. 76, nos 1-3, , p. 75–87 (ISSN 0303-2647, DOI 10.1016/j.biosystems.2004.05.030)
- ↑ Thomas C. G. Bosch, Alexander Klimovich, Tomislav Domazet-Lošo et Stefan Gründer, « Back to the Basics: Cnidarians Start to Fire », Trends in neurosciences, vol. 40, no 2, , p. 92–105 (ISSN 0166-2236, PMID 28041633, PMCID PMC5285349, DOI 10.1016/j.tins.2016.11.005, lire en ligne, consulté le )
- ↑ (en) Ruppert, E.E., Fox, R.S. et Barnes, R.D., Invertebrate Zoology : a functional evolutionary approach, Belmont (Calif.), Brooks / Cole, , 7e éd., 989 p. (ISBN 0-03-025982-7)
- ↑ « Aspects de la nutrition des coraux (scléractiniaires) », sur vieoceane.free.fr.
- ↑ « Description des cnidaires », sur univ-lehavre.fr.
- ↑ Jacqueline Goy, « Les paradoxes des méduses », Pour la Science, no 299, , p. 39 (résumé)
- ↑ World Register of Marine Species, consulté le 5 février 2018
- ↑ Integrated Taxonomic Information System (ITIS), www.itis.gov, CC0 https://doi.org/10.5066/F7KH0KBK, consulté le 20 février 2014
- ↑ [Marques & Collins 2004] Antonio Carlos Marques et Allen G. Collins, « Cladistic analysis of Medusozoa and cnidarian evolution », Invertebrate Biology, vol. 123, no 1, , p. 23-42 (lire en ligne [sur researchgate.net], consulté en ).
- ↑ Halanych, K.M., « The New View of Animal Phylogeny », Annual Review of Ecology, Evolution, and Systematics, vol. 35, , p. 229–256 (DOI 10.1146/annurev.ecolsys.35.112202.130124, lire en ligne [archive du ], consulté le )
- ↑ Borchiellini, C., Manuel, M., Alivon, E., Boury-Esnault, N., Vacelet J. et Le Parco, Y., « Sponge paraphyly and the origin of Metazoa », Journal of Evolutionary Biology, vol. 14, no 1, , p. 171–179 (PMID 29280585, DOI 10.1046/j.1420-9101.2001.00244.x
 , S2CID 25119754)
, S2CID 25119754)
- ↑ Medina, M., Collins, A.G., Silberman, J.D. et Sogin, M.L., « Evaluating hypotheses of basal animal phylogeny using complete sequences of large and small subunit rRNA », Proceedings of the National Academy of Sciences, vol. 98, no 17, , p. 9707–9712 (PMID 11504944, PMCID 55517, DOI 10.1073/pnas.171316998 , Bibcode 2001PNAS...98.9707M)
- ↑ Müller, W.E.G., Li, J., Schröder, H.C., Qiao, L. et Wang, X., « The unique skeleton of siliceous sponges (Porifera; Hexactinellida and Demospongiae) that evolved first from the Urmetazoa during the Proterozoic: a review », Biogeosciences, vol. 4, no 2, , p. 219–232 (DOI 10.5194/bg-4-219-2007 , Bibcode 2007BGeo....4..219M, lire en ligne)
- Collins, A.G., « Phylogeny of Medusozoa and the Evolution of Cnidarian Life Cycles », Journal of Evolutionary Biology, vol. 15, no 3, , p. 418–432 (DOI 10.1046/j.1420-9101.2002.00403.x , S2CID 11108911)
- ↑ Wallberg, A., Thollesson, M., Farris, J.S. et Jondelius, U., « The phylogenetic position of the comb jellies (Ctenophora) and the importance of taxonomic sampling », Cladistics, vol. 20, no 6, , p. 558–578 (PMID 34892961, DOI 10.1111/j.1096-0031.2004.00041.x, S2CID 86185156)
- ↑ Marques, A.C. et Collins, A.G., « Cladistic analysis of Medusozoa and cnidarian evolution », Invertebrate Biology, vol. 123, no 1, , p. 23–42 (DOI 10.1111/j.1744-7410.2004.tb00139.x, lire en ligne, consulté le )
- ↑ (en) K. Seipel et V. Schmid, « Evolution of striated muscle: Jellyfish and the origin of triploblasty », Developmental Biology, vol. 282, no 1, , p. 14–26 (PMID 15936326, DOI 10.1016/j.ydbio.2005.03.032 )
- ↑ Miller, D.J., Ball, E.E. et Technau, U., « Cnidarians and ancestral genetic complexity in the animal kingdom », Trends in Genetics, vol. 21, no 10, , p. 536–539 (PMID 16098631, DOI 10.1016/j.tig.2005.08.002)
- ↑ Technau, U., Rudd, S. et Maxwell, P, « Maintenance of ancestral complexity and non-metazoan genes in two basal cnidarians », Trends in Genetics, vol. 21, no 12, , p. 633–639 (PMID 16226338, DOI 10.1016/j.tig.2005.09.007)
- ↑ D. R. Smith, E. Kayal, A. A. Yanagihara, A. G. Collins, S. Pirro et P. J. Keeling, « First Complete Mitochondrial Genome Sequence from a Box Jellyfish Reveals a Highly Fragmented Linear Architecture and Insights into Telomere Evolution », Genome Biology and Evolution, vol. 4, no 1, , p. 52–58 (PMID 22117085, PMCID 3268669, DOI 10.1093/gbe/evr127)