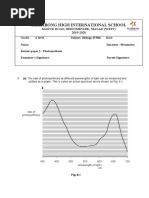
Ascent of Sap: 3 Theories: (A) Godlewski S Relay-Pump Theory
Ascent of Sap: 3 Theories: (A) Godlewski S Relay-Pump Theory
Download as pdf or txt
You might also like
- Polymorphism in CoelenteratesDocument30 pagesPolymorphism in Coelenteratessrisankaraglobalacademy7No ratings yet
- Grade 9 Biology Term II Paper 2 MCQDocument16 pagesGrade 9 Biology Term II Paper 2 MCQRahulBansuman100% (1)
- Aibítir: The Irish Alphabet in Botanical Art CatalogueDocument49 pagesAibítir: The Irish Alphabet in Botanical Art CatalogueJane Stark100% (3)
- 4.4 Ascent of Sap and TheoriesDocument16 pages4.4 Ascent of Sap and Theoriesankurbiology75% (4)
- Water: The Origin of Life from Water to the Molecules of LifeFrom EverandWater: The Origin of Life from Water to the Molecules of LifeNo ratings yet
- FalgDocument101 pagesFalgshaikh85No ratings yet
- Photoperiodism: 1. Short Day PlantsDocument6 pagesPhotoperiodism: 1. Short Day PlantsKarm JiNo ratings yet
- Magneto Hydro Dynamic Generators: A Solution For Future Energy CrisisDocument7 pagesMagneto Hydro Dynamic Generators: A Solution For Future Energy CrisisZipronNo ratings yet
- Ohmic Heating of FoodsDocument18 pagesOhmic Heating of FoodsHarish SrinivasanNo ratings yet
- Bubble-Size Dependence of The Critical Electrolyte Concentration For Inhibition of CoalescenceDocument8 pagesBubble-Size Dependence of The Critical Electrolyte Concentration For Inhibition of CoalescenceJose Luis Barrientos RiosNo ratings yet
- ''Traditional Arabic and Islamic Medicine''Document4 pages''Traditional Arabic and Islamic Medicine''tandraini100% (1)
- Duckweed FactsheetDocument3 pagesDuckweed FactsheetRocky SarmientoNo ratings yet
- Transport in Plant - Class NotesDocument32 pagesTransport in Plant - Class Noteszaijah taylor4ANo ratings yet
- Edward LINACRE - Airdrop Irrigation SystemDocument4 pagesEdward LINACRE - Airdrop Irrigation Systemfafsdfdf12100% (1)
- Eden Blue Gold Garden BlendDocument87 pagesEden Blue Gold Garden BlendEden Blue GoldNo ratings yet
- Part 1: Water Potential and Components: CBCS/ Plant Physiology/ Botany (H) /sem V LecturesDocument14 pagesPart 1: Water Potential and Components: CBCS/ Plant Physiology/ Botany (H) /sem V LecturesK S ChatterjeeNo ratings yet
- Transport in Plants NotesDocument28 pagesTransport in Plants NotesYashpal Suwansia100% (1)
- Plant Tissue SystemsDocument3 pagesPlant Tissue Systemselizabeth shaw gonzalez100% (1)
- Clay Pot Refrigerator PDFDocument6 pagesClay Pot Refrigerator PDFAvinash RaghooNo ratings yet
- Plant Physiolog-WPS OfficeDocument45 pagesPlant Physiolog-WPS OfficeShubhendu Chattopadhyay100% (1)
- Biodynamic AgricultureDocument3 pagesBiodynamic AgricultureEliane MasonNo ratings yet
- Synthesis of FerrofluidDocument3 pagesSynthesis of FerrofluidValentin-AngeloUzunovNo ratings yet
- Angiosperms Lab ActivityDocument11 pagesAngiosperms Lab ActivityHaris Khan100% (1)
- Mass Transport Processes in The Electro-Oxidation of Organic Compounds For Wastewater TreatmentDocument8 pagesMass Transport Processes in The Electro-Oxidation of Organic Compounds For Wastewater Treatmentprasad wakodeNo ratings yet
- Principles and Methods of Pollen Analysis, Pollen Production, Liberation, Dispersal, Deposition and PreservationDocument13 pagesPrinciples and Methods of Pollen Analysis, Pollen Production, Liberation, Dispersal, Deposition and PreservationAbdulkarim AcidriNo ratings yet
- Glauber Salt For Energy StorageDocument45 pagesGlauber Salt For Energy StoragearjmandquestNo ratings yet
- Respiration in ArthropodaDocument36 pagesRespiration in Arthropodadark.phoenix91124No ratings yet
- Bio 102 Lab ReportDocument9 pagesBio 102 Lab ReportLisa IvesterNo ratings yet
- Photosynthesis LabDocument4 pagesPhotosynthesis LabSanzida TaslimNo ratings yet
- Tritium: Approved For Public Release Distribution UnlimitedDocument40 pagesTritium: Approved For Public Release Distribution UnlimitedIngenioerenNo ratings yet
- Water Fuel CellDocument2 pagesWater Fuel Cellronachaif3191100% (1)
- Cdi T1 2018-19 G11 TGDocument149 pagesCdi T1 2018-19 G11 TGShadi Abbas50% (2)
- Echolocation in AnimalsDocument3 pagesEcholocation in AnimalschristinaNo ratings yet
- Effer VesDocument4 pagesEffer VesFarkas RoskoNo ratings yet
- Pollen Wall - ExtraexinousWallMaterialsDocument44 pagesPollen Wall - ExtraexinousWallMaterialsSUDHA GUPTA100% (1)
- Femto-H2 Transmuttation Proves The Current Nucleus Model IncorrectDocument9 pagesFemto-H2 Transmuttation Proves The Current Nucleus Model IncorrectInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Royal College of Science Narowal: CyanobacteriaDocument11 pagesRoyal College of Science Narowal: Cyanobacteriakiran maqsoodNo ratings yet
- Manual Electricty and MagnetismDocument13 pagesManual Electricty and MagnetismDennis Cosmod100% (1)
- ECOLOGY & Climatic FactorsDocument17 pagesECOLOGY & Climatic Factorsrajshgupta01No ratings yet
- Plant Physiology and Biochemistry: Photosynthesis and Transport of Organic SubstancesDocument38 pagesPlant Physiology and Biochemistry: Photosynthesis and Transport of Organic SubstancesAsh AliNo ratings yet
- The Neurohistological TechniquesDocument4 pagesThe Neurohistological Techniquesapi-3846255100% (2)
- THEORIES OF SHOOT APICAL MERISTEM NotesDocument11 pagesTHEORIES OF SHOOT APICAL MERISTEM NotesAbhimanyu Pandey100% (2)
- Biological InvasionDocument9 pagesBiological InvasionHarsh Vardhan SinghNo ratings yet
- Telluric Radiation Antenna - Articles Amp French Patent 641575Document12 pagesTelluric Radiation Antenna - Articles Amp French Patent 641575qqqqNo ratings yet
- Decarboxylation Wagner ADocument24 pagesDecarboxylation Wagner AEnrico MedianoNo ratings yet
- V. Kaznacheev - Kozyrev MirrorsDocument28 pagesV. Kaznacheev - Kozyrev MirrorsKalpa235No ratings yet
- Timer Temporary Immersion Bio-Reactor (Basic DIY Diagram)Document1 pageTimer Temporary Immersion Bio-Reactor (Basic DIY Diagram)potadoNo ratings yet
- Experimental Comparative Analysis of Clay Pot Refrigeration Using Two Different Designs of PotsDocument6 pagesExperimental Comparative Analysis of Clay Pot Refrigeration Using Two Different Designs of PotsInternational Journal of Latest Research in Engineering and TechnologyNo ratings yet
- Immediate download Living Rainbow H2O 1st Edition Mae-Wan Ho ebooks 2024Document81 pagesImmediate download Living Rainbow H2O 1st Edition Mae-Wan Ho ebooks 2024mpelleivens100% (2)
- Anatomical Structure of RootDocument5 pagesAnatomical Structure of RootkingNo ratings yet
- Artificial Generation of Accustic and Gravitational Waves in The AtmosphereDocument10 pagesArtificial Generation of Accustic and Gravitational Waves in The AtmosphereNOLINKNEWSNo ratings yet
- Anwita Jha Magh 4, 2077Document25 pagesAnwita Jha Magh 4, 2077Anwita JhaNo ratings yet
- BIO ProjectDocument17 pagesBIO ProjectHârsh V ÎshwãkårmāNo ratings yet
- ORMUS 2 of 3Document7 pagesORMUS 2 of 3TimjoboNo ratings yet
- Plasma Physics: Fundamentals ofDocument7 pagesPlasma Physics: Fundamentals ofVipul JoshiNo ratings yet
- The National Institute of Engineering.: Tesla CoilDocument7 pagesThe National Institute of Engineering.: Tesla Coilnav NNo ratings yet
- 03 Marincic Civric MilovanovicDocument18 pages03 Marincic Civric MilovanovicThami KNo ratings yet
- 4.animalkingdom ResonanceDocument80 pages4.animalkingdom ResonanceEkta ManglaniNo ratings yet
- Mechanism of Absorption of WaterDocument6 pagesMechanism of Absorption of WaterketakeeNo ratings yet
- All NCERT SCIENTISTS by Seep PahujaDocument36 pagesAll NCERT SCIENTISTS by Seep PahujaLakshmiNo ratings yet
- Review of Related LiteratureDocument10 pagesReview of Related LiteratureRobert Kier Tanquerido Tomaro100% (1)
- Nuclear Alternative: Redesigning Our Model of the Structure of MatterFrom EverandNuclear Alternative: Redesigning Our Model of the Structure of MatterNo ratings yet
- CBSE Grade 10 Biology QP Dated 02.07.2021Document1 pageCBSE Grade 10 Biology QP Dated 02.07.2021RahulBansumanNo ratings yet
- CBSE Grade 10 Biology QP Dated 10.7.2020 KEYDocument5 pagesCBSE Grade 10 Biology QP Dated 10.7.2020 KEYRahulBansumanNo ratings yet
- Grade 9 Biology Term II Paper 6 ATPDocument11 pagesGrade 9 Biology Term II Paper 6 ATPRahulBansumanNo ratings yet
- 0610 w18 QP 21-CIE-IGCSE-BiologyDocument20 pages0610 w18 QP 21-CIE-IGCSE-BiologyRahulBansuman100% (1)
- 00 6th N-Guide MPCB Term-1 Mcs IndexDocument1 page00 6th N-Guide MPCB Term-1 Mcs IndexRahulBansumanNo ratings yet
- Cambridge International AS and A Level Biology: Paper 3 - Advanced Practical SkillsDocument36 pagesCambridge International AS and A Level Biology: Paper 3 - Advanced Practical SkillsRahulBansuman100% (1)
- June 2017 (v2) QP - Paper 4 CIE Biology IGCSEDocument20 pagesJune 2017 (v2) QP - Paper 4 CIE Biology IGCSERahulBansumanNo ratings yet
- Co-Ordination in HumansDocument14 pagesCo-Ordination in HumansRahulBansumanNo ratings yet
- A Level Review Paper 2Document10 pagesA Level Review Paper 2RahulBansumanNo ratings yet
- History of DurianDocument16 pagesHistory of DurianMarc Nixon PelayoNo ratings yet
- S5 - Q2 - Summative Test 3Document4 pagesS5 - Q2 - Summative Test 3Jason PlazaNo ratings yet
- Land Management Willow Tree FactsheetDocument4 pagesLand Management Willow Tree FactsheetnorthernmeldrewNo ratings yet
- Padhle 10th - How Do Organisms Reproduce + Integrated PYQs (1) - CompressedDocument9 pagesPadhle 10th - How Do Organisms Reproduce + Integrated PYQs (1) - CompressedkarrmanbhatiaNo ratings yet
- KD ScheduleDocument162 pagesKD Schedulethomas_hanNo ratings yet
- Ethnobotany - WikipediaDocument4 pagesEthnobotany - WikipediasaxophonemanNo ratings yet
- The Magic Garden Class 3 Notes CBSE English Chapter 1 (PDF)Document4 pagesThe Magic Garden Class 3 Notes CBSE English Chapter 1 (PDF)Chaitra CyNo ratings yet
- Plant Tissues: PBOT 111: Lecture 5Document42 pagesPlant Tissues: PBOT 111: Lecture 5Cham RosarioNo ratings yet
- Lab Report1Document13 pagesLab Report1pratiwi kusumaNo ratings yet
- MCQ - L1-Characteristics & Classification of Living Organisms (Multiple Choice) 1 QPDocument18 pagesMCQ - L1-Characteristics & Classification of Living Organisms (Multiple Choice) 1 QPkaveesh_93No ratings yet
- Bankable ProjectDocument39 pagesBankable ProjectpradeepNo ratings yet
- 2013 Rivals Et Al Dietary Ecology of Extant Guanaco (Lama Guanicoe) From Southern PatagoniaDocument10 pages2013 Rivals Et Al Dietary Ecology of Extant Guanaco (Lama Guanicoe) From Southern PatagoniaRicardoDennisMezaNiflaNo ratings yet
- Physiological Disorders in Cole Vegetable CropsDocument33 pagesPhysiological Disorders in Cole Vegetable CropsAkash DesaiNo ratings yet
- Handbook8 Fruit - And.vegetables 1Document21 pagesHandbook8 Fruit - And.vegetables 1Gaurav GhoshNo ratings yet
- Symbiosis Laboratory Investigation 1Document3 pagesSymbiosis Laboratory Investigation 1api-282243542No ratings yet
- Processing: Fig. 16. Mature Luffa Gourd FruitsDocument1 pageProcessing: Fig. 16. Mature Luffa Gourd FruitsanothersomeguyNo ratings yet
- Lupang HinirangDocument12 pagesLupang HinirangYsabelle Yu YagoNo ratings yet
- BS - EN - 351!1!2007 Durability of WoodDocument24 pagesBS - EN - 351!1!2007 Durability of WoodGerardo BeckerNo ratings yet
- Don Bosco Splendid Home 1 Terminal Examination-2017 Class-VIII BiologyDocument3 pagesDon Bosco Splendid Home 1 Terminal Examination-2017 Class-VIII BiologyproodootNo ratings yet
- Form 3 Chapter 4Document7 pagesForm 3 Chapter 4naza9775100% (7)
- Agripreneurship Project: Litchicana-Litchi Cultivation UnitDocument37 pagesAgripreneurship Project: Litchicana-Litchi Cultivation UnitcentralresourcesgmaiNo ratings yet
- Exer 3Document7 pagesExer 3Adonis MiñanoNo ratings yet
- Drink in The Wild - Teas, Cordials, Jams and More (2003)Document137 pagesDrink in The Wild - Teas, Cordials, Jams and More (2003)Alberto González100% (2)
- Heuchera and Heucherella For The GardenDocument15 pagesHeuchera and Heucherella For The GardenDennis CareyNo ratings yet
- Plas Moly SisDocument1 pagePlas Moly SisRaymund BondeNo ratings yet
- PhytoremediationDocument42 pagesPhytoremediationSeroKeretaMasaroWidiarNo ratings yet
- Journal Issaas v15n1 POSTER ABSTRACTSDocument28 pagesJournal Issaas v15n1 POSTER ABSTRACTSChai YawatNo ratings yet
- The Botanic Garden. Erasmus DarwinDocument474 pagesThe Botanic Garden. Erasmus DarwinnuilaNo ratings yet
- Plant Tissue CultureDocument71 pagesPlant Tissue CultureShree Krishna Adhikari100% (6)