The Cell Cycle & Apoptosis
The Cell Cycle & Apoptosis
Download as pdf or txt
At a glance
Powered by AI
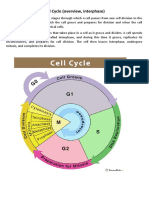
The key takeaways are that the cell cycle consists of 4 main phases (G1, S, G2, M) and is tightly regulated by various proteins like cyclins, CDKs, and CDK inhibitors to control cell growth and division.
The main phases of the cell cycle are G1, S, G2, and M. G1 is the growth phase, S is DNA synthesis, G2 is more growth, and M is mitosis where the cell divides.
The main proteins that regulate and control the cell cycle are the cyclins (especially cyclin D and E), cyclin-dependent kinases (CDK4, CDK6, CDK2), and CDK inhibitors (p16, p21, p27). Tumor suppressor proteins like Rb and p53 also play important regulatory roles.
You might also like
- Snorks Activity RevisedDocument8 pagesSnorks Activity RevisedFatima Viterbo Montalvo100% (1)
- Molecular Cell Biology Lodish 6th Edition Test BankDocument6 pagesMolecular Cell Biology Lodish 6th Edition Test BankCharles BlairNo ratings yet
- TranslationDocument78 pagesTranslationdrmukhtiarbaigNo ratings yet
- Genetics, Lecture 11 (Lecture Notes)Document16 pagesGenetics, Lecture 11 (Lecture Notes)Ali Al-QudsiNo ratings yet
- Control of Prokaryotic (Bacterial) Genes: AP BiologyDocument15 pagesControl of Prokaryotic (Bacterial) Genes: AP BiologysilNo ratings yet
- Apoptosis and CancerDocument20 pagesApoptosis and CancerSajjad Ahmad0% (1)
- 3 LipidsDocument44 pages3 LipidsSuresh ChovatiyaNo ratings yet
- Cell Signaling in Space and Time:: Where Proteins Come Together and When They're ApartDocument20 pagesCell Signaling in Space and Time:: Where Proteins Come Together and When They're ApartHumbertoNo ratings yet
- Nucleus: Click To Edit Master Subtitle StyleDocument29 pagesNucleus: Click To Edit Master Subtitle StyleAzifah ZakariaNo ratings yet
- The Citric Acid CycleDocument43 pagesThe Citric Acid CycleMohammed AlMujainiNo ratings yet
- Chapter 13Document56 pagesChapter 13Hậu VũNo ratings yet
- Overview of Cell Cycle by Javali.GDocument15 pagesOverview of Cell Cycle by Javali.GJavali.GNo ratings yet
- Module-1: Genomics and Proteomics NotesDocument8 pagesModule-1: Genomics and Proteomics NotesNishtha KhannaNo ratings yet
- Cell CommunicationDocument25 pagesCell Communicationprehealthhelp100% (12)
- Molecular Biology of The Cell Cumulative Final Exam Study GuideDocument22 pagesMolecular Biology of The Cell Cumulative Final Exam Study Guiderazgriz1211No ratings yet
- Chromatin RemodellingDocument234 pagesChromatin Remodellingplastioid4079No ratings yet
- Protein TransportDocument13 pagesProtein TransportvmshanesNo ratings yet
- Cell Signalling Notes Merit Life Sciences1Document61 pagesCell Signalling Notes Merit Life Sciences1merit tutorials com NET JRF JAM TUTORIALSNo ratings yet
- Ch11 - Cell Signaling SlidesDocument36 pagesCh11 - Cell Signaling SlidesValine Cysteine MethionineNo ratings yet
- 1.transcription 2012Document80 pages1.transcription 2012Fajar AgNo ratings yet
- Cell Biology and Membrane BiochemistryDocument106 pagesCell Biology and Membrane BiochemistryVijay Bhasker TekulapallyNo ratings yet
- Restriction Enzymes MSC BiotechDocument37 pagesRestriction Enzymes MSC BiotechRoneet Ghosh0% (1)
- Cell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemDocument70 pagesCell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemGovinda BiswasNo ratings yet
- Intracellular SignallingDocument45 pagesIntracellular Signallingsana iqbalNo ratings yet
- Isolation of Genomic DNA From Animal TissuesDocument6 pagesIsolation of Genomic DNA From Animal TissuesSanmugam SubbaramaniamNo ratings yet
- RIBOZYMESDocument17 pagesRIBOZYMESSharan GayathrinathanNo ratings yet
- Chloroplast and PhotosynthesisDocument19 pagesChloroplast and PhotosynthesisFizul HelmiNo ratings yet
- Physiology of Muscle ContractionDocument3 pagesPhysiology of Muscle ContractionAJ AbraganNo ratings yet
- 002 Cell Signalling PathwaysDocument138 pages002 Cell Signalling PathwaysstarykNo ratings yet
- Respiratory Chain & Oxidative PhosphorylationDocument57 pagesRespiratory Chain & Oxidative PhosphorylationHanifa AffianiNo ratings yet
- The Cell Cycle: For Campbell Biology, Ninth EditionDocument98 pagesThe Cell Cycle: For Campbell Biology, Ninth Edition46bwilsonNo ratings yet
- Molecular BiologyDocument9 pagesMolecular Biologyrifat RasheedNo ratings yet
- Cell Signalling 1.2 2017Document61 pagesCell Signalling 1.2 2017PutriNo ratings yet
- Genome Organization in ProkaryotesDocument8 pagesGenome Organization in ProkaryotesVijay Kishore75% (4)
- CH 11 PPT Cell Communication 1Document77 pagesCH 11 PPT Cell Communication 1api-270681964No ratings yet
- ReplicationDocument45 pagesReplicationAleena MustafaNo ratings yet
- 36 ch10 PDFDocument12 pages36 ch10 PDFMajid Al-hachami100% (1)
- Affinity ChromatographyDocument3 pagesAffinity ChromatographyNithya Ram100% (1)
- Cell Communication - DADocument66 pagesCell Communication - DAMuhammad Ardiansyah MNo ratings yet
- Regulation of Gene ExpressionDocument5 pagesRegulation of Gene ExpressionHanumat SinghNo ratings yet
- Biochemistry - Final Exam Biology 020.305 December 15, 2011 Instructions For ExaminationDocument11 pagesBiochemistry - Final Exam Biology 020.305 December 15, 2011 Instructions For ExaminationJenna ReynoldsNo ratings yet
- Protein Synthesis (Transcription and Translation) : Dr. Marhaen Hardjo, M.Biomed, PHDDocument159 pagesProtein Synthesis (Transcription and Translation) : Dr. Marhaen Hardjo, M.Biomed, PHDsNo ratings yet
- Second Messengers Camp CGMPDocument42 pagesSecond Messengers Camp CGMPMirza Shaharyar BaigNo ratings yet
- G-Protein Coupled ReceptorsDocument8 pagesG-Protein Coupled ReceptorsHyunji KimNo ratings yet
- Western BlottingDocument13 pagesWestern BlottingAshfaq Fazal100% (1)
- Endomembrane SystemDocument26 pagesEndomembrane SystemPreeti SainiNo ratings yet
- COURSE WORK MOLECULAR BIOLOGY & GeneticsDocument3 pagesCOURSE WORK MOLECULAR BIOLOGY & Geneticsusaeed00000No ratings yet
- Intra Cellular Vesicular TrafficDocument48 pagesIntra Cellular Vesicular TrafficKanishkaNo ratings yet
- Oxidative PhosphorylationDocument33 pagesOxidative PhosphorylationJithendra Babu0% (1)
- Biotin in Metabolism and Its Relationship To Human DiseaseDocument9 pagesBiotin in Metabolism and Its Relationship To Human DiseaseMaikel Ignacio Z. PeñaNo ratings yet
- Regulation of Cell CycleDocument5 pagesRegulation of Cell CyclemohsinNo ratings yet
- Size Exclusion Column ChromatographyDocument8 pagesSize Exclusion Column ChromatographySmeetha Kaur100% (1)
- Lysosomes PresentationDocument53 pagesLysosomes PresentationCaroline Stephenson50% (2)
- Cells Study Guide 2020 KEYDocument4 pagesCells Study Guide 2020 KEYLaarni GeeNo ratings yet
- Ultrastructure Nucleus: BiologyDocument14 pagesUltrastructure Nucleus: BiologyAshu ShewaleNo ratings yet
- Apoptosis by Dr. Sanjiv Kumar-1Document27 pagesApoptosis by Dr. Sanjiv Kumar-1चौधरी हरिओम सौरोतNo ratings yet
- DNA ReplicationDocument32 pagesDNA ReplicationSudipta MandolNo ratings yet
- BIOL223-Lab 7Document40 pagesBIOL223-Lab 7chicken fries100% (2)
- Regulation of Gene Expression in Prokaryotic and Eukaryotic CellsDocument13 pagesRegulation of Gene Expression in Prokaryotic and Eukaryotic CellsMifta KhuljannahNo ratings yet
- Cell Division MitosisDocument37 pagesCell Division MitosisAshley PerezNo ratings yet
- BIO109 Mitosis Vs Meiosis Assignment: NameDocument1 pageBIO109 Mitosis Vs Meiosis Assignment: NameCindy TestaNo ratings yet
- AP Biology SupplementDocument86 pagesAP Biology SupplementLama DiriyehNo ratings yet
- Cellular Reproduction: How To Preserve Genetic InformationDocument45 pagesCellular Reproduction: How To Preserve Genetic InformationAditya RachmadNo ratings yet
- Concepts of Genetics 2nd Edition by Brooker ISBN Test BankDocument24 pagesConcepts of Genetics 2nd Edition by Brooker ISBN Test Bankpamela100% (39)
- HANDOUT - KEY Photosynthesis and Cellular Respiration Review Sheet 2017Document5 pagesHANDOUT - KEY Photosynthesis and Cellular Respiration Review Sheet 2017hannahwawarofskyNo ratings yet
- MYP 4 IDU Energy RESPIRATION 1Document82 pagesMYP 4 IDU Energy RESPIRATION 1aseda.stanhopeessamuahp6No ratings yet
- Bioorganic & Medicinal Chemistry LettersDocument4 pagesBioorganic & Medicinal Chemistry LettersSana MazharNo ratings yet
- MitochondriaDocument14 pagesMitochondriaPhoebe KimNo ratings yet
- Assignment 1 - Gene Regulation - Edpuzzle Video With QuestionsDocument3 pagesAssignment 1 - Gene Regulation - Edpuzzle Video With QuestionsThuy DinhNo ratings yet
- Mutation Chromosome Change in Number and StructureDocument53 pagesMutation Chromosome Change in Number and StructureyukbhuviNo ratings yet
- Onion Mitosis LabDocument8 pagesOnion Mitosis LabTonyDiazNo ratings yet
- Cambridge Assessment International Education: Biology 9700/41 May/June 2018Document17 pagesCambridge Assessment International Education: Biology 9700/41 May/June 2018shabanaNo ratings yet
- 153C Final Exam Study Guide-2Document6 pages153C Final Exam Study Guide-2Soji AdimulaNo ratings yet
- Transcription and TranslationDocument58 pagesTranscription and Translationkevin_ramos007No ratings yet
- B2.2 NotesDocument13 pagesB2.2 NotesteddyenNo ratings yet
- Maria ShelloDocument16 pagesMaria ShelloMaria Shello SaruedaNo ratings yet
- Carbohydrate MetabolismDocument83 pagesCarbohydrate Metabolismapi-3826907100% (7)
- Chapter 5-Mitochondria and Aerobic Respiration - 複本Document32 pagesChapter 5-Mitochondria and Aerobic Respiration - 複本Wai Kwong ChiuNo ratings yet
- Pengantar MetabolismeDocument19 pagesPengantar MetabolismeHildya KusmiNo ratings yet
- Cell CycleDocument10 pagesCell CycleDrexel DalaygonNo ratings yet
- Bioenergetics and MetabolismDocument56 pagesBioenergetics and MetabolismAww AddNo ratings yet
- Overview of Biochemical EndocrinologyDocument20 pagesOverview of Biochemical EndocrinologyGauri KashyapNo ratings yet
- Biological Oxidation by SatishDocument12 pagesBiological Oxidation by SatishBiochemistry DenNo ratings yet
- HHMI Eukaryotic Cell Cycle and CancerDocument3 pagesHHMI Eukaryotic Cell Cycle and Cancerhopkins.ian0618No ratings yet
- Molecular Basis of Inheritance NotesDocument12 pagesMolecular Basis of Inheritance NotesAmritpal Singh100% (2)
- Partoza - JB, Concept Article - Endomembranesystem - DocxDocument4 pagesPartoza - JB, Concept Article - Endomembranesystem - DocxJenny PartozaNo ratings yet
- Q2 SHS - PT - Gen Biology 1 - 2022 - 23Document6 pagesQ2 SHS - PT - Gen Biology 1 - 2022 - 23Diane GuilaranNo ratings yet
- AP Biology Exploration Guide - Cell Metabolism #3Document6 pagesAP Biology Exploration Guide - Cell Metabolism #3Gunnar OlsonNo ratings yet