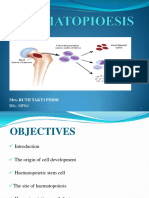
IUC - Haematology - Lecture 2 - Hemopoiesis-2021
IUC - Haematology - Lecture 2 - Hemopoiesis-2021
Download as pptx, pdf, or txt
You might also like
- Shaukat Khanum Memorial Cancer Hospital & Research Centre: SPC Nature: SPC SiteDocument3 pagesShaukat Khanum Memorial Cancer Hospital & Research Centre: SPC Nature: SPC SiteMisum AliNo ratings yet
- AFIP Manual New HaemDocument76 pagesAFIP Manual New HaemDr.A SHAHID SiddiquiNo ratings yet
- Haematopoiesis: DR Rosline Hassan Hematology Department School of Medical Sciences Universiti Sains MalaysiaDocument46 pagesHaematopoiesis: DR Rosline Hassan Hematology Department School of Medical Sciences Universiti Sains Malaysialow_sernNo ratings yet
- ASE SC 1 Animal & Plant Cell ColoringDocument3 pagesASE SC 1 Animal & Plant Cell ColoringAlfie Cabario100% (1)
- Hemopoiesis PDFDocument14 pagesHemopoiesis PDFrysnawahyu13No ratings yet
- Exercise 5 HematopoiesisDocument11 pagesExercise 5 HematopoiesisAngela AninangNo ratings yet
- Hematopoiesis: Pluri - Several) or Hemocytoblasts. These Cells Have The Capacity To Develop Into Many Different Types ofDocument12 pagesHematopoiesis: Pluri - Several) or Hemocytoblasts. These Cells Have The Capacity To Develop Into Many Different Types ofRajender ArutlaNo ratings yet
- Bio 2 Assignment 3Document16 pagesBio 2 Assignment 3Arun RajNo ratings yet
- Lym, Throm, GranuDocument12 pagesLym, Throm, Granumaxwell amponsahNo ratings yet
- HaematopoiesisDocument77 pagesHaematopoiesisFrancis PrathyushaNo ratings yet
- BSC Lecture Notes Semester 1Document7 pagesBSC Lecture Notes Semester 1satasha28No ratings yet
- Erythropoiesis: Presented by DR Etu-Efeotor T. PDocument88 pagesErythropoiesis: Presented by DR Etu-Efeotor T. PPrincewill SeiyefaNo ratings yet
- CELLS and ORGANS of IMMUNE SYSTEM - FormattedDocument26 pagesCELLS and ORGANS of IMMUNE SYSTEM - FormattedSantiago MuñozNo ratings yet
- Hematopoietic DevelopmentDocument6 pagesHematopoietic DevelopmentSea ClestNo ratings yet
- Hematopoiesisby VarishaDocument5 pagesHematopoiesisby VarishaJoão CanjimuenaNo ratings yet
- غير معروف Introduction to haematology-1 (Muhadharaty)Document47 pagesغير معروف Introduction to haematology-1 (Muhadharaty)aliabumrfghNo ratings yet
- Haematopoiesis JonesDocument15 pagesHaematopoiesis Joneskatemabenjamin53No ratings yet
- Immunology Unit 1Document14 pagesImmunology Unit 1a.parameshwari1234No ratings yet
- Blood Formation LectureDocument24 pagesBlood Formation Lecturehassan aryaniNo ratings yet
- Hemopoisis NotesDocument4 pagesHemopoisis NotesIqra AsifNo ratings yet
- TranslateDocument6 pagesTranslatenazarovmuhammad1131No ratings yet
- NOTESDocument101 pagesNOTESPaul AshburnerNo ratings yet
- Haematopoitik System: Prof - Dr. Adi Koesoema Aman SPPK (KH) - Dr. Malayana Nasution Mked - SPPKDocument43 pagesHaematopoitik System: Prof - Dr. Adi Koesoema Aman SPPK (KH) - Dr. Malayana Nasution Mked - SPPKrubyniNo ratings yet
- RBC Lecture Outline Erythropoiesis 2010-01-02Document5 pagesRBC Lecture Outline Erythropoiesis 2010-01-02anil awaseNo ratings yet
- D 11 53 5f0f108789ad7Document34 pagesD 11 53 5f0f108789ad7jsbart330No ratings yet
- Heamtopoises ReviewerDocument17 pagesHeamtopoises ReviewerClyde BaltazarNo ratings yet
- Unit 02 Hematopoiesis and Normal Leukocytes FunctionDocument54 pagesUnit 02 Hematopoiesis and Normal Leukocytes Functionusamah.a.sayedNo ratings yet
- Haematopoiesis: Haematopoiesis (from Ancient Greek: αἷμα, "blood"; ποιεῖν "to make") (or hematopoiesis in theDocument5 pagesHaematopoiesis: Haematopoiesis (from Ancient Greek: αἷμα, "blood"; ποιεῖν "to make") (or hematopoiesis in theDivakaran SathyamurthyNo ratings yet
- 01 - Bone Marrow & Hematopoiesis - DR Najeeb HematologyDocument8 pages01 - Bone Marrow & Hematopoiesis - DR Najeeb Hematologyhiba jasim100% (1)
- Hema TransesDocument25 pagesHema TransesNikoh Anthony EwayanNo ratings yet
- The Formed Elements of Blood Have Relatively Short LifespansDocument14 pagesThe Formed Elements of Blood Have Relatively Short LifespansBilly James EncalladoNo ratings yet
- HEMOPOIESISDocument26 pagesHEMOPOIESISinkwellinsights23No ratings yet
- Leucopoiesis and ThrombopoiesisDocument6 pagesLeucopoiesis and Thrombopoiesishardworkeffiong14No ratings yet
- Chapter 20: The Cardiovascular System: Blood: Embryology Atlas HematopoiesisDocument8 pagesChapter 20: The Cardiovascular System: Blood: Embryology Atlas Hematopoiesisfjestefania6892No ratings yet
- LECTURE 2 - HEMATOPOIESIS and ERYTHROPOIESIS - 10 - 17 - 2020Document49 pagesLECTURE 2 - HEMATOPOIESIS and ERYTHROPOIESIS - 10 - 17 - 2020apoorva krishnagiriNo ratings yet
- Approach To Hematologic DisordersDocument8 pagesApproach To Hematologic DisorderstyasNo ratings yet
- Bone Marrow: Andy Mclarnon, University of Birmingham, UkDocument2 pagesBone Marrow: Andy Mclarnon, University of Birmingham, Uknavita guptaNo ratings yet
- ErythropoiesisDocument3 pagesErythropoiesisSubhasish BarikNo ratings yet
- Hematopoeitic System& Blood, KBK 2015 LDLDocument80 pagesHematopoeitic System& Blood, KBK 2015 LDLgita dwi ananda100% (1)
- Haematopoiesis, Gfs and Requirements Dr.tugu 17-10-23Document42 pagesHaematopoiesis, Gfs and Requirements Dr.tugu 17-10-23ataya2442No ratings yet
- Erythropoiesis WORDDocument10 pagesErythropoiesis WORDANIRBAN ASUTOSH SWAINNo ratings yet
- Formation of Blood CellsDocument20 pagesFormation of Blood CellsEbong Michael100% (1)
- Development and Maintenance of Hematopoietic Tissues: Colony-Forming Units (Cfus)Document13 pagesDevelopment and Maintenance of Hematopoietic Tissues: Colony-Forming Units (Cfus)Tyler KingNo ratings yet
- Hematology Review NotesDocument28 pagesHematology Review NotesMeLaiyee89% (9)
- Semeiology of Hematopoietic SystemDocument18 pagesSemeiology of Hematopoietic SystemIvanNo ratings yet
- Topic 1Document21 pagesTopic 1Ria AccadNo ratings yet
- Bone MarrowDocument3 pagesBone Marrowhk9976614No ratings yet
- Erythropoiesis: Definition Site of ErythropoiesisDocument6 pagesErythropoiesis: Definition Site of ErythropoiesisAnusuya SNo ratings yet
- Cell DifferentiationDocument16 pagesCell DifferentiationHarika KeshavNo ratings yet
- Hematopoiesis New 161214142246Document29 pagesHematopoiesis New 161214142246Vikram KumarNo ratings yet
- Hematology NotesDocument24 pagesHematology Notesddrei13No ratings yet
- Stem CellDocument8 pagesStem CellHanumat SinghNo ratings yet
- CLINPATH-01.Hematopoiesis & Morphology of Blood CellsDocument10 pagesCLINPATH-01.Hematopoiesis & Morphology of Blood CellsCharisse Angelica MacedaNo ratings yet
- He Ma To PoiesisDocument64 pagesHe Ma To Poiesismatthew deguzman100% (1)
- Hema Report PDFDocument25 pagesHema Report PDFdaliaNo ratings yet
- Chapter1-S 20-21Document36 pagesChapter1-S 20-21Shoo ShaaNo ratings yet
- CL1 HaemopoiesisDocument41 pagesCL1 HaemopoiesisSamuel George100% (1)
- Lecture 3 (1) HaematopoeisisDocument20 pagesLecture 3 (1) Haematopoeisismaxwell amponsahNo ratings yet
- Histologi HematopoeitikDocument29 pagesHistologi HematopoeitikFadhil AhmadNo ratings yet
- Hematology GNDocument94 pagesHematology GNJan Lian Enriquez NoolNo ratings yet
- Blood SciencesDocument47 pagesBlood Sciencescaroline vaughanNo ratings yet
- 4.1.1 Draw and Label A Diagram of A Motor Unit.: Movement AnalysisDocument30 pages4.1.1 Draw and Label A Diagram of A Motor Unit.: Movement AnalysisChristyle AquinoNo ratings yet
- XI CH - 06 Anatomy of Flowering Plants - 220605 - 233239Document17 pagesXI CH - 06 Anatomy of Flowering Plants - 220605 - 233239Annu BaijalNo ratings yet
- The Digestive System 3Document4 pagesThe Digestive System 3geniusgurl91No ratings yet
- Module 8. Animal Organ SystemDocument151 pagesModule 8. Animal Organ SystemMarkAnthony PagaNo ratings yet
- Blood ReservationDocument1 pageBlood Reservationfritzie dara f adjilaniNo ratings yet
- MIDTERM EXAMINATION IN GEN BIO 1 Cell TheoryDocument4 pagesMIDTERM EXAMINATION IN GEN BIO 1 Cell TheoryIAN MAR VALENZUELA100% (1)
- Tissues & MembranesDocument9 pagesTissues & MembranesKrystel Morales DizonNo ratings yet
- Histopathology Unit: PretestDocument3 pagesHistopathology Unit: PretestMary CabalceNo ratings yet
- Introduction To Professional PracticeDocument95 pagesIntroduction To Professional PracticeRevista PongNo ratings yet
- Blood TypingDocument3 pagesBlood Typingangalee garuti0% (1)
- Jawapan BioDocument9 pagesJawapan BioJesus SmithNo ratings yet
- Cell Organelles FunctionsDocument1 pageCell Organelles FunctionsFedomessi10 Is a geniusNo ratings yet
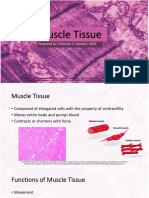
- Muscle TissueDocument40 pagesMuscle TissuefsdafvNo ratings yet
- Anatomy & Physiology The Muscular System Flashcards - QuizletDocument5 pagesAnatomy & Physiology The Muscular System Flashcards - Quizletmalenya1No ratings yet
- Acid-Fast StainDocument1 pageAcid-Fast StainMerve Nur GülerNo ratings yet
- Quipper 1 - Earth and Life Science - CellDocument17 pagesQuipper 1 - Earth and Life Science - CellPhil Christian MerinoNo ratings yet
- Components of Prokaryotic Cells: Eral Structure of A Prokaryotic Cell: This Figure Shows The Generalized Structure ofDocument3 pagesComponents of Prokaryotic Cells: Eral Structure of A Prokaryotic Cell: This Figure Shows The Generalized Structure ofJess MCDNo ratings yet
- Chapter 19Document42 pagesChapter 19Dr. Manish KumarNo ratings yet
- Lect 1 Introduction To Histo PathologyDocument14 pagesLect 1 Introduction To Histo PathologyAbdul Hafeez100% (2)
- Histology and CytologyDocument4 pagesHistology and CytologyDharshini KumarNo ratings yet
- Cell MembraneDocument5 pagesCell MembraneJenica CruzNo ratings yet
- Histology of The EyeDocument5 pagesHistology of The EyeshindyNo ratings yet
- Biomechanics Unit 3Document24 pagesBiomechanics Unit 3RajuNo ratings yet
- B2.2 NotesDocument13 pagesB2.2 NotesteddyenNo ratings yet
- Defining The Practice of Medical TechnologyDocument47 pagesDefining The Practice of Medical TechnologyCharles Ian OliquinoNo ratings yet
- Cell Biology & Organisation: Body/what-Cells-In-The-Human-Body-Live-The-LongestDocument17 pagesCell Biology & Organisation: Body/what-Cells-In-The-Human-Body-Live-The-LongestDewi SallehNo ratings yet
- Science Art Integrated Project 10thDocument8 pagesScience Art Integrated Project 10thravmeetkaurchpNo ratings yet
- Chapter 8 - Musculoskeletal SystemDocument9 pagesChapter 8 - Musculoskeletal SystemIbby HooriyaNo ratings yet