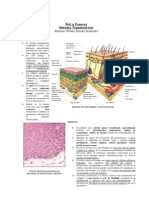
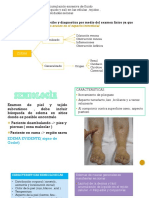
Fisiopatologia Del Cancer
Fisiopatologia Del Cancer
Descargar como docx, pdf o txt
También podría gustarte
- El Sistema LocomotorDocumento2 páginasEl Sistema Locomotormapi196493% (15)
- Trast. Alimentacion PDFDocumento87 páginasTrast. Alimentacion PDFtroleira100% (1)
- Loteria ITSDocumento10 páginasLoteria ITSKaren Díaz100% (3)
- Equipo de Protección Individual para Uso en Planta de Tratamiento de Aguas ResidualesDocumento6 páginasEquipo de Protección Individual para Uso en Planta de Tratamiento de Aguas ResidualesJuan Manuel Cruz67% (3)
- Patologia de Huesos y ArticulacionesDocumento9 páginasPatologia de Huesos y ArticulacionesMARIA FERNANDA TOBAR MUNOZAún no hay calificaciones
- Clase 9. Lesiones LigamentosasDocumento5 páginasClase 9. Lesiones LigamentosasALEJANDRAAún no hay calificaciones
- Cancer GastricoDocumento23 páginasCancer GastricoRicardo Andres Avila Ocampo0% (1)
- Enfermedades Del Aparato LocomotorDocumento34 páginasEnfermedades Del Aparato LocomotorRafael N NietoAún no hay calificaciones
- Puntos GatilloDocumento14 páginasPuntos GatilloBernardita BelénAún no hay calificaciones
- Clasificacion Diverticulitis TablasDocumento4 páginasClasificacion Diverticulitis TablasRenato PomarAún no hay calificaciones
- Teoría Páncreas Endocrino - MEHU 632Documento26 páginasTeoría Páncreas Endocrino - MEHU 632CRISTEL GRACIELA HUAMAN DELGADOAún no hay calificaciones
- 2.toma de Ecg Mod 2018Documento28 páginas2.toma de Ecg Mod 2018esmos20105826Aún no hay calificaciones
- Cambios Anatómicos y Fisiológicos en El Sistema Cardiovascular y Respiratorio en El Adulto MayorDocumento14 páginasCambios Anatómicos y Fisiológicos en El Sistema Cardiovascular y Respiratorio en El Adulto MayorJosé Lucero HuamanAún no hay calificaciones
- Esofagitis Por Causticos PDFDocumento34 páginasEsofagitis Por Causticos PDFAna Quispe CuroAún no hay calificaciones
- Tratamiento Fisioterapéutico en La Tendinopatia Del SupraespinosoDocumento24 páginasTratamiento Fisioterapéutico en La Tendinopatia Del SupraespinosokaisoongAún no hay calificaciones
- Historia Clinica Final (2) (1) DR JOEDocumento23 páginasHistoria Clinica Final (2) (1) DR JOEclavel777Aún no hay calificaciones
- Alteraciones Del PeneDocumento8 páginasAlteraciones Del Peneyakelin100% (1)
- Programa de Prevención y Control IAASDocumento37 páginasPrograma de Prevención y Control IAASFrancisca BartschAún no hay calificaciones
- Medicina de AlturaDocumento15 páginasMedicina de AlturaFernando Rios ZeladaAún no hay calificaciones
- Espondilitis AnquilosanteDocumento35 páginasEspondilitis AnquilosanteArmando Contreras Sandoval100% (1)
- Monografía Sobre Enfermedades Que Afectan Al Esófago. Enfermería PAF USS.Documento38 páginasMonografía Sobre Enfermedades Que Afectan Al Esófago. Enfermería PAF USS.Jose Maria Castillo PlazaAún no hay calificaciones
- CASO Clinico EpocDocumento22 páginasCASO Clinico EpocMonse OroscoAún no hay calificaciones
- Investigación Documental Del CannabisDocumento30 páginasInvestigación Documental Del CannabisAlejandraAún no hay calificaciones
- LUMBOCIATICADocumento6 páginasLUMBOCIATICApopeludoAún no hay calificaciones
- Patologia EstomatologicaDocumento62 páginasPatologia EstomatologicalizAún no hay calificaciones
- ¿Qué Es La Anosognosia PDFDocumento8 páginas¿Qué Es La Anosognosia PDFgrace prambsAún no hay calificaciones
- Ojos y OidosDocumento58 páginasOjos y OidosAngie DiazAún no hay calificaciones
- Examen Físico Aparato Genital Femenino Capt 95 GuarderasDocumento11 páginasExamen Físico Aparato Genital Femenino Capt 95 GuarderasCristian ZumarragaAún no hay calificaciones
- Fisiología EndocrinaDocumento9 páginasFisiología EndocrinaNay OrtizAún no hay calificaciones
- Enfermedades OseasDocumento5 páginasEnfermedades OseasFabio Ribera SaavedraAún no hay calificaciones
- Sesion de Salud - Lavado de ManosDocumento14 páginasSesion de Salud - Lavado de ManosVivas AndréAún no hay calificaciones
- TCSC Tejido Linfatico DR Paul Upao 2023Documento28 páginasTCSC Tejido Linfatico DR Paul Upao 2023GUSTAVO ANTONIO SANCHEZ MASIASAún no hay calificaciones
- Musc. Esqueletico I-ColumnaDocumento35 páginasMusc. Esqueletico I-ColumnaLaura López Del Castillo LalydelcaAún no hay calificaciones
- Tendinopatias PDFDocumento3 páginasTendinopatias PDFRodrigo SánchezAún no hay calificaciones
- Guion Microbiologia Medica IDocumento69 páginasGuion Microbiologia Medica IIadpa BarquisimetoAún no hay calificaciones
- Lesiones Del Manguito RotadorDocumento29 páginasLesiones Del Manguito RotadorJuan DLGAún no hay calificaciones
- Clase Cardiovascular SíntomasDocumento57 páginasClase Cardiovascular SíntomasVerty100% (1)
- Aines ActualizadoDocumento47 páginasAines ActualizadoCamila Gómez MejíaAún no hay calificaciones
- LinfangiomasDocumento38 páginasLinfangiomasBarbara Helen Stanley100% (1)
- Ruidos Cardiacos PatologicosDocumento32 páginasRuidos Cardiacos PatologicosValeria FabrisAún no hay calificaciones
- Estratificación de Riesgo de Enfermedad Coronaria Con MétodosDocumento22 páginasEstratificación de Riesgo de Enfermedad Coronaria Con MétodosAshley CguizarAún no hay calificaciones
- Introducción A La Semiología MedicaDocumento7 páginasIntroducción A La Semiología MedicaSherlock HolmesAún no hay calificaciones
- 3ro Piel y FanerasDocumento4 páginas3ro Piel y FaneraskcodonerAún no hay calificaciones
- Penicilina MODIDocumento28 páginasPenicilina MODIivan menaAún no hay calificaciones
- Valvulopatias PulmonaresDocumento19 páginasValvulopatias PulmonaresYessenia Castro RodriguezAún no hay calificaciones
- Fisiopatologia EsofagoDocumento102 páginasFisiopatologia EsofagoNicte CamachoAún no hay calificaciones
- HemiplejiaDocumento7 páginasHemiplejiaAdel VasquezAún no hay calificaciones
- RESPIDocumento8 páginasRESPIAmpa Ro DriguezAún no hay calificaciones
- Neuroftalmologia 01X02Documento5 páginasNeuroftalmologia 01X02Katherine ReyesAún no hay calificaciones
- PROYECTODocumento35 páginasPROYECTODEISY MANAUREAún no hay calificaciones
- Bronquitis CronicaDocumento2 páginasBronquitis CronicaMila MillaAún no hay calificaciones
- Rehabilitación Respiratoria 2020-IDocumento52 páginasRehabilitación Respiratoria 2020-ILunaNuevaAún no hay calificaciones
- Sistema GastrointestinalDocumento49 páginasSistema Gastrointestinaljhon saicoAún no hay calificaciones
- Esclerosis Sistémica ProgresivaDocumento36 páginasEsclerosis Sistémica ProgresivaPANKEKASAún no hay calificaciones
- EspondilodicitisDocumento4 páginasEspondilodicitisJose VRAún no hay calificaciones
- EsofagitisDocumento106 páginasEsofagitisClaudia Mercedes Achacollo AchaAún no hay calificaciones
- Exploración de Piel y Faneras y Sistema LinfaticoDocumento13 páginasExploración de Piel y Faneras y Sistema LinfaticocancerosoAún no hay calificaciones
- Tronco EncefalicoDocumento5 páginasTronco Encefalicopepepearl11Aún no hay calificaciones
- Informe EPOCDocumento10 páginasInforme EPOCweolasAún no hay calificaciones
- Cancer Del EstomagoDocumento9 páginasCancer Del EstomagoDelinaAún no hay calificaciones
- AnemiaDocumento8 páginasAnemiaKatherynIzquierdoReynaAún no hay calificaciones
- Monografia D Oncogenes y ProtooncogenesDocumento11 páginasMonografia D Oncogenes y ProtooncogenesCesar Huallpa0% (1)
- Genes y Cáncer IDocumento7 páginasGenes y Cáncer IHernán UrbanoAún no hay calificaciones
- Drogas AntianginosasDocumento38 páginasDrogas AntianginosasEnrique Domingo Pol OrdoñezAún no hay calificaciones
- Sepsis UrinariaDocumento10 páginasSepsis UrinariaEnrique Domingo Pol OrdoñezAún no hay calificaciones
- Temas de Nutricion Basica PDFDocumento225 páginasTemas de Nutricion Basica PDFTomenossAún no hay calificaciones
- Temas de Nutricion Basica PDFDocumento225 páginasTemas de Nutricion Basica PDFTomenossAún no hay calificaciones
- PADI MADRID. (Programa de Atención Dental Infantil para La Comunidad de Madrid)Documento11 páginasPADI MADRID. (Programa de Atención Dental Infantil para La Comunidad de Madrid)nanoAún no hay calificaciones
- EdemaDocumento9 páginasEdemaCarmen Ramirez QuiñonezAún no hay calificaciones
- Introducción A La Psicologia SocialDocumento5 páginasIntroducción A La Psicologia SocialMiktlan PerezAún no hay calificaciones
- Aparato DigestivoDocumento18 páginasAparato DigestivoVane Jth100% (1)
- Pelajes Equinos Genetica y TransmicionDocumento55 páginasPelajes Equinos Genetica y TransmicionFlorencia MioriAún no hay calificaciones
- Caso 4Documento1 páginaCaso 4Sarai Rosales0% (1)
- Valoracion Recien NacidoDocumento56 páginasValoracion Recien NacidoJuan Carlos Flores MejíaAún no hay calificaciones
- Contactos Dentarios Durante La MasticaciónDocumento9 páginasContactos Dentarios Durante La Masticacióncamilo pallesAún no hay calificaciones
- Cilindros UrinariosDocumento15 páginasCilindros UrinariosMagdaly Correa100% (1)
- Cólera Porcina (Bartolo Moreno)Documento17 páginasCólera Porcina (Bartolo Moreno)Fredrick Alvarado67% (3)
- Anatomia Comparada Aparato CirculatorioDocumento9 páginasAnatomia Comparada Aparato CirculatorioAnghela YubetsyAún no hay calificaciones
- Colorantes SinteticosDocumento15 páginasColorantes SinteticosJuan Jose Huarca Sutta100% (1)
- TESIS - Ciencia, Realidad y Metodo en La Obra de Linus Pauling +++++++Documento468 páginasTESIS - Ciencia, Realidad y Metodo en La Obra de Linus Pauling +++++++larg140884646Aún no hay calificaciones
- Vitamin AsDocumento39 páginasVitamin AsBrenda MAún no hay calificaciones
- FisiologíaDocumento30 páginasFisiologíaFernanda Rodríguez0% (1)
- Clase08.2010 Ruptura Del Receso Invernal. Uso de Cianamida Hidrogenada PDFDocumento4 páginasClase08.2010 Ruptura Del Receso Invernal. Uso de Cianamida Hidrogenada PDFAriana PuiconAún no hay calificaciones
- CEPRE ONLINE2.0 - BIOLOGÍA - Clases Gratuitas 09052020Documento4 páginasCEPRE ONLINE2.0 - BIOLOGÍA - Clases Gratuitas 09052020Hugo FloresAún no hay calificaciones
- Confort HumanoDocumento5 páginasConfort HumanoyadiraaromAún no hay calificaciones
- Minsur - Seguridad y SaludDocumento9 páginasMinsur - Seguridad y Saludjhimmy ronaldAún no hay calificaciones
- Control Prenatal ENEO-UNAMDocumento17 páginasControl Prenatal ENEO-UNAMNidia Mendoza Q100% (1)
- Plantas AntigripalesDocumento4 páginasPlantas AntigripalesZuley Geraldine Hurtado ArceAún no hay calificaciones
- Preguntas Regulacion de La Expresion GenicaDocumento3 páginasPreguntas Regulacion de La Expresion GenicaHelen Romero100% (3)
- Sistema Endocrino y DigestivoDocumento13 páginasSistema Endocrino y DigestivoLuis DazaAún no hay calificaciones
- Conceptualizar Los Problemas Del ClienteDocumento15 páginasConceptualizar Los Problemas Del ClienteAlex DiesAún no hay calificaciones
- Placenta PreviaDocumento5 páginasPlacenta PreviaFelipe ZapataAún no hay calificaciones
- Aparato RespiratorioDocumento22 páginasAparato RespiratorioAndrea MagdalenaAún no hay calificaciones