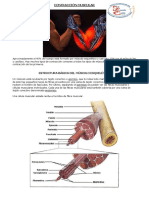
CAPITULOS
CAPITULOS
Descargar como pdf o txt
También podría gustarte
- Anatomia y Fisiología del Sistema Nervioso IIDe EverandAnatomia y Fisiología del Sistema Nervioso IICalificación: 4 de 5 estrellas4/5 (28)
- Anatomía y Fisiología del Sistema NerviosoDe EverandAnatomía y Fisiología del Sistema NerviosoCalificación: 4 de 5 estrellas4/5 (19)
- Capítulo 6 Contracción Del Músculo EsqueléticoDocumento13 páginasCapítulo 6 Contracción Del Músculo EsqueléticoDiego José Castro CastañoAún no hay calificaciones
- UNIDAD II. Capítulo 6 - Contracción Del Músculo Esquelético - Ana Montejo, FisiologíaDocumento25 páginasUNIDAD II. Capítulo 6 - Contracción Del Músculo Esquelético - Ana Montejo, FisiologíaIrma SánchezAún no hay calificaciones
- Contracción Muscular PDFDocumento6 páginasContracción Muscular PDFDani Morales DellatorreAún no hay calificaciones
- 3.1 Excitabilidad - Músculo EsqueléticoDocumento16 páginas3.1 Excitabilidad - Músculo Esqueléticogochita1504Aún no hay calificaciones
- Repaso FisiologiaDocumento17 páginasRepaso FisiologiaSALOME PANTOJAAún no hay calificaciones
- Contracción Del Musculo Esqueletico - FisiologíaDocumento8 páginasContracción Del Musculo Esqueletico - FisiologíaDara DosAún no hay calificaciones
- Resumen HistologiaDocumento34 páginasResumen HistologiavalentinaAún no hay calificaciones
- FISIOLOGIA GUYTON UNIDAD 1 SEMANA 2 Capitulo 6Documento2 páginasFISIOLOGIA GUYTON UNIDAD 1 SEMANA 2 Capitulo 6LUCIA DEL CARMEN OTOYA CORONELAún no hay calificaciones
- Cap. 6Documento30 páginasCap. 6RafaelaAún no hay calificaciones
- Capitulo 6 Guyton PDFDocumento35 páginasCapitulo 6 Guyton PDFRodrigo CañeteAún no hay calificaciones
- Fisiologia de Guyton y Hall Tratado de Fisiologia MedicaDocumento5 páginasFisiologia de Guyton y Hall Tratado de Fisiologia MedicaMa FerAún no hay calificaciones
- FISIOLOGÍA MUSCULAR EdzsDocumento38 páginasFISIOLOGÍA MUSCULAR EdzsCieloNaupariCalderónAún no hay calificaciones
- Contracción Del Músculo EsqueléticoDocumento58 páginasContracción Del Músculo EsqueléticoAnnália GestaAún no hay calificaciones
- Fisiología de La Contracción Del Músculo EsqueléticoDocumento17 páginasFisiología de La Contracción Del Músculo Esqueléticovikybonilla938Aún no hay calificaciones
- Tema 19 FHDocumento14 páginasTema 19 FHMaria MingAún no hay calificaciones
- Manual FisiologiaDocumento12 páginasManual Fisiologianiliram.floresAún no hay calificaciones
- Control Muscular Del Movimiento PDFDocumento33 páginasControl Muscular Del Movimiento PDFNicolas Diaz100% (1)
- Cap 6-Contraccion de Músculo EsqueléticoDocumento13 páginasCap 6-Contraccion de Músculo EsqueléticoDawidh Ortiz GarciaAún no hay calificaciones
- Cap 6 Contraccion Del Musculo Esqueletico 1 MaryDocumento36 páginasCap 6 Contraccion Del Musculo Esqueletico 1 MaryLaura VelasquezAún no hay calificaciones
- 7 Observacion de Celulas Musculares PDFDocumento37 páginas7 Observacion de Celulas Musculares PDFmartin mahomaAún no hay calificaciones
- Tema 12 - Los MusculosDocumento11 páginasTema 12 - Los Musculoszahrazahoudi23Aún no hay calificaciones
- Unidad 17 Biología Celular Sistema Nervioso - Muscular - InmuneDocumento101 páginasUnidad 17 Biología Celular Sistema Nervioso - Muscular - Inmunemartingandolfo1Aún no hay calificaciones
- Sistema MotorDocumento10 páginasSistema Motoreilj00120Aún no hay calificaciones
- Fisiología de La Contracción Muscular.2018Documento21 páginasFisiología de La Contracción Muscular.2018Freddy Ademar Plata MamaniAún no hay calificaciones
- Tema 8: Tejido Muscular: - MiofibrillasDocumento8 páginasTema 8: Tejido Muscular: - Miofibrillascarlota martinezAún no hay calificaciones
- Neurofisiología IV PDFDocumento29 páginasNeurofisiología IV PDFClaudia S. HernándezAún no hay calificaciones
- Contracción Del Musculo EsqueléticoDocumento12 páginasContracción Del Musculo EsqueléticoEfren MoraAún no hay calificaciones
- OrganizaciónMuscular 31 Marzo2020Documento34 páginasOrganizaciónMuscular 31 Marzo2020luanebutterflyAún no hay calificaciones
- Bases Biofísicas Del ElectrocardiogramaDocumento15 páginasBases Biofísicas Del Electrocardiogramaneomartin666Aún no hay calificaciones
- Tema 5 FisioDocumento5 páginasTema 5 FisiouniversejddAún no hay calificaciones
- Fisiología Muscular - MEHU 438Documento35 páginasFisiología Muscular - MEHU 438CIELO NATALY RIOJAS DE LA CRUZAún no hay calificaciones
- Tejido MuscularDocumento65 páginasTejido Muscularnatalyconstanzacabrera123Aún no hay calificaciones
- Sistema MuscularDocumento55 páginasSistema Muscularjose luis navarreteAún no hay calificaciones
- FisiologíaDocumento24 páginasFisiologíajoaquinAún no hay calificaciones
- Cap 6 Contracción Del Músculo EsqueléticoDocumento9 páginasCap 6 Contracción Del Músculo Esqueléticorodrigo4590Aún no hay calificaciones
- Ebook - Biologia PDFDocumento40 páginasEbook - Biologia PDFcassianedejesusxrAún no hay calificaciones
- Tejido Muscular - @aprendermedicinaDocumento5 páginasTejido Muscular - @aprendermedicinashmarsowluAún no hay calificaciones
- Clase 8 Tejido Muscular 1Documento68 páginasClase 8 Tejido Muscular 1Karen CunalataAún no hay calificaciones
- Músculo EsqueleticoDocumento61 páginasMúsculo EsqueleticoPaola Rocha100% (1)
- Fisiologia de La Contraccion MuscularDocumento30 páginasFisiologia de La Contraccion Muscularchiquillintivo100% (1)
- Clase 8 Tejido Muscular 1Documento52 páginasClase 8 Tejido Muscular 1bryanjhoan32Aún no hay calificaciones
- Expo. Fibra Muscular - GuionDocumento5 páginasExpo. Fibra Muscular - GuioncaroAún no hay calificaciones
- Fisio MuscularDocumento45 páginasFisio MuscularYami YamAún no hay calificaciones
- TejidomuscularDocumento52 páginasTejidomuscularMartina La PortaAún no hay calificaciones
- Fisiología - Módulo 6Documento8 páginasFisiología - Módulo 6Santiago RodriguezAún no hay calificaciones
- Tejido MuscularDocumento4 páginasTejido MuscularCamila Valdés AlvearAún no hay calificaciones
- Tejido MuscularDocumento53 páginasTejido MuscularUSUARIO LUCILAAún no hay calificaciones
- Contracción Del Músculo EsqueléticoDocumento7 páginasContracción Del Músculo EsqueléticoJaneth Serrano ArmentaAún no hay calificaciones
- Tejidos MuscularesDocumento45 páginasTejidos MuscularesHéctor MorenoAún no hay calificaciones
- Unidad 3 resumenDocumento19 páginasUnidad 3 resumenlorenzorutilrAún no hay calificaciones
- Contracción MuscularDocumento24 páginasContracción MuscularheleanyAún no hay calificaciones
- 06 Biología de La Contracción MuscularDocumento32 páginas06 Biología de La Contracción MuscularCamila ArguelloAún no hay calificaciones
- MECANISMO GENERAL y MOLECULAR DE LA CONTRACCIÓNDocumento7 páginasMECANISMO GENERAL y MOLECULAR DE LA CONTRACCIÓNKatiAriasAún no hay calificaciones
- Cap. 06 Contracción Del Músculo EsqueléticoDocumento6 páginasCap. 06 Contracción Del Músculo EsqueléticoJocelyne Benito100% (1)
- Tejido Muscular Esquelético 2020Documento48 páginasTejido Muscular Esquelético 2020Marcos DíasAún no hay calificaciones
- Mecanismo de Contracción MuscularDocumento17 páginasMecanismo de Contracción MuscularDanielly RibeiroAún no hay calificaciones
- Histología y Embrología del Sitema NerviosoDe EverandHistología y Embrología del Sitema NerviosoCalificación: 4 de 5 estrellas4/5 (4)
- Anatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2De EverandAnatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2Aún no hay calificaciones
- Taller No. 1 Tejido EpitelialDocumento11 páginasTaller No. 1 Tejido Epitelialangelly pinillaAún no hay calificaciones
- Determinación de Variante DuDocumento5 páginasDeterminación de Variante DuAntonioAún no hay calificaciones
- Resumen AnemiasDocumento8 páginasResumen AnemiasMar ArredondoAún no hay calificaciones
- BioloDocumento7 páginasBioloCarmen Diez RufoAún no hay calificaciones
- Aguinaldo MargariteñoDocumento6 páginasAguinaldo MargariteñoAngel ContrerasAún no hay calificaciones
- ZoologíaDocumento3 páginasZoologíaivan thomasAún no hay calificaciones
- Biología Molecular Y CelularDocumento4 páginasBiología Molecular Y CelularMarielen MotaAún no hay calificaciones
- ReportDocumento3 páginasReportAndres SáezAún no hay calificaciones
- Hongos Imperfectos o DeuteromycotaDocumento3 páginasHongos Imperfectos o DeuteromycotaMarjorie Lizeth Tineo DíazAún no hay calificaciones
- Módulo Biología 3ro Riquelme 2022Documento97 páginasMódulo Biología 3ro Riquelme 2022Yamila AbadAún no hay calificaciones
- MitocondriaDocumento26 páginasMitocondriaMarleny Agudelo GracianoAún no hay calificaciones
- StimulateDocumento4 páginasStimulateJOSE LUIS ALATORREAún no hay calificaciones
- Problemas examenes PSICOBIOLOGÍA UNEDDocumento6 páginasProblemas examenes PSICOBIOLOGÍA UNEDBlanca EstevaAún no hay calificaciones
- Cuadro Comparativo MicrobiologiaDocumento4 páginasCuadro Comparativo MicrobiologiaEduardoAún no hay calificaciones
- INFORME - S2 - Aislamiento de DNA PlasmidicoDocumento4 páginasINFORME - S2 - Aislamiento de DNA PlasmidicoPaulina Lucrecia Barajas HernandezAún no hay calificaciones
- F-AC-21 TALLER DE CLASE P13 9 2021 Genetica MendelianaDocumento2 páginasF-AC-21 TALLER DE CLASE P13 9 2021 Genetica MendelianaIvan Uzumaki0% (1)
- Herencia MitocondrialDocumento14 páginasHerencia MitocondrialDiego Perez leteAún no hay calificaciones
- Ciencia y Tecnologia Martes 05Documento9 páginasCiencia y Tecnologia Martes 05FreddyChiroque AncajimaAún no hay calificaciones
- Exam 1erparc BiologíaDocumento2 páginasExam 1erparc Biologíajesus MendozaAún no hay calificaciones
- Sílabus Productos Biologicos MI 505 - 2023-IDocumento17 páginasSílabus Productos Biologicos MI 505 - 2023-INathaly Jovana Contreras HerreraAún no hay calificaciones
- Taller Excepciones de Las Leyes de MendelDocumento3 páginasTaller Excepciones de Las Leyes de MendelRafael SeguraAún no hay calificaciones
- El Tigre y Los ValoresDocumento6 páginasEl Tigre y Los ValoresCanerox De PereiraAún no hay calificaciones
- Taller 2. Regulaciòn o Control de La Expresiòn GenicaDocumento9 páginasTaller 2. Regulaciòn o Control de La Expresiòn GenicaDaryl TinjacaAún no hay calificaciones
- Laboratorio 6Documento4 páginasLaboratorio 6kiwik8554Aún no hay calificaciones
- Práctica 1 Biología IIDocumento4 páginasPráctica 1 Biología IImedinaulloafanyAún no hay calificaciones
- Taller 1 BiotecnologiaDocumento2 páginasTaller 1 BiotecnologiaMaicol CastellanosAún no hay calificaciones
- Biologia Semana 1Documento22 páginasBiologia Semana 1Leoniza Sanchez AlvarezAún no hay calificaciones
- Tema 3-ANATOMIA Y FISIOLOGIA DE LA PIELDocumento33 páginasTema 3-ANATOMIA Y FISIOLOGIA DE LA PIELLupen MadridAún no hay calificaciones
- Actividad de Aprendizaje N°05-Ct-4to-2024.Documento6 páginasActividad de Aprendizaje N°05-Ct-4to-2024.antonype73Aún no hay calificaciones
- Resumen II Bloque - MicroDocumento182 páginasResumen II Bloque - Microgabriela loarcaAún no hay calificaciones