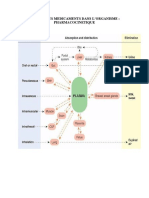
3 - Metabolisme Des Protides
3 - Metabolisme Des Protides
Télécharger au format pdf ou txt
Vous aimerez peut-être aussi
- QuestionsDocument15 pagesQuestionsHajar ChadlaouiPas encore d'évaluation
- Rapport Pig Farming Rwanda Invest BE v4Document23 pagesRapport Pig Farming Rwanda Invest BE v4Portier FabienPas encore d'évaluation
- Metabolisme Des Acides AminesDocument9 pagesMetabolisme Des Acides AminesHea FPas encore d'évaluation
- Métabolisme Des Acide AminésDocument26 pagesMétabolisme Des Acide Aminés[AE]100% (4)
- Biologie CellulaireDocument24 pagesBiologie CellulairealmnaouarPas encore d'évaluation
- Les Acides Et Les BasesDocument3 pagesLes Acides Et Les BasesRemy HebertPas encore d'évaluation
- Bio Cell À ProjeterDocument51 pagesBio Cell À ProjeterSalauwma GenievaPas encore d'évaluation
- Metabolisme Des Lipides 2011 2012Document13 pagesMetabolisme Des Lipides 2011 2012Ka TiaPas encore d'évaluation
- Métabolisme Des GlucidesDocument28 pagesMétabolisme Des GlucidesilaPas encore d'évaluation
- Student-NUTRITION CARBONEE-2021-2022Document99 pagesStudent-NUTRITION CARBONEE-2021-2022IDTH EBM ApplicationPas encore d'évaluation
- Acides Aminés Pour Aller Plus LoinDocument15 pagesAcides Aminés Pour Aller Plus LoinCooper WaynePas encore d'évaluation
- Cours3 ProteinesDocument28 pagesCours3 ProteinesLinda KoundziPas encore d'évaluation
- TD N1Géréralités Sur Les Enzymes 2022Document20 pagesTD N1Géréralités Sur Les Enzymes 2022Sarah AitelhadjPas encore d'évaluation
- FLAvonoideDocument99 pagesFLAvonoideFatima Zahra ElayyoubiPas encore d'évaluation
- Sulfamides HypoglycemiantsDocument12 pagesSulfamides Hypoglycemiantsfarracygmailcom100% (1)
- COURS - 3 - Procaryote EucaryoteDocument40 pagesCOURS - 3 - Procaryote EucaryoteNESRINE NABIPas encore d'évaluation
- Chap 1 Les Glucides MedecineDocument34 pagesChap 1 Les Glucides MedecineManel GadouchePas encore d'évaluation
- Drogues À Anthocyanosides Et À CoumarinesDocument8 pagesDrogues À Anthocyanosides Et À CoumarinesZied BouabenePas encore d'évaluation
- Les DiurétiquesDocument26 pagesLes DiurétiquesBMA-medecinePas encore d'évaluation
- BiochimieDocument109 pagesBiochimieedgardokafPas encore d'évaluation
- Chapitre 1 La PharmacocinétiqueDocument5 pagesChapitre 1 La PharmacocinétiquebilalPas encore d'évaluation
- Metabo Partie2Document6 pagesMetabo Partie2la biochimiste100% (2)
- UE2 La-Membrane-PlasmiqueDocument17 pagesUE2 La-Membrane-Plasmiquesun-nee-chan9Pas encore d'évaluation
- Materiel Genetique PDFDocument25 pagesMateriel Genetique PDFSoubere Mahamoud100% (1)
- DR A. AouicheDocument43 pagesDR A. Aouicheamira laggounPas encore d'évaluation
- Biochimie Structurale 2Document105 pagesBiochimie Structurale 2PekoJhames08gmail.comPas encore d'évaluation
- ProteinesDocument13 pagesProteinesyaya camaraPas encore d'évaluation
- Métabolisme Du GlycogèneDocument7 pagesMétabolisme Du GlycogèneaxelPas encore d'évaluation
- Peptides Et ProtéinesDocument12 pagesPeptides Et ProtéinesIngenieur AgroPas encore d'évaluation
- Glucides 1Document47 pagesGlucides 1Rania MaddahPas encore d'évaluation
- Cycle CellulaireDocument37 pagesCycle CellulairegigiPas encore d'évaluation
- 3 Membrane PlasmiqueDocument53 pages3 Membrane PlasmiqueLina RazekPas encore d'évaluation
- Métabolisme Des GlucidesDocument6 pagesMétabolisme Des GlucidesHouda SoniaPas encore d'évaluation
- EndocrinologieDocument46 pagesEndocrinologieAhmedou EllaPas encore d'évaluation
- Bioenerg Enzymo 2023Document91 pagesBioenerg Enzymo 2023Ghars Adam100% (1)
- Nutrition VegetauxDocument5 pagesNutrition VegetauxnacerbensaPas encore d'évaluation
- Cours 5 Systeme Nerveux AutonomeDocument80 pagesCours 5 Systeme Nerveux AutonomeSofyane AboulkacemPas encore d'évaluation
- Microbiologie Générale DR TOUAITIADocument35 pagesMicrobiologie Générale DR TOUAITIAعالمي الجميلPas encore d'évaluation
- 01 - La Communication IntercellulaireDocument8 pages01 - La Communication IntercellulaireHadil BenslamaPas encore d'évaluation
- Notes Phytochimie - RésuméDocument5 pagesNotes Phytochimie - RésumémimiPas encore d'évaluation
- La Physiologie VégétaleDocument15 pagesLa Physiologie VégétaleradhiamecroPas encore d'évaluation
- 11-Structure Et Fonction Des ACIDES AMINESDocument55 pages11-Structure Et Fonction Des ACIDES AMINESBook Hunter100% (2)
- Infectiologie Cours 28 Les Virus OncogenesDocument18 pagesInfectiologie Cours 28 Les Virus OncogenesPelle100% (1)
- C1 VirologieDocument18 pagesC1 VirologieSABRINA AYOUZPas encore d'évaluation
- BOTAanatomieDocument10 pagesBOTAanatomieNsangwaPas encore d'évaluation
- ExercicesDocument44 pagesExercicesmanal treegreen100% (1)
- Polycopie Cours Bioch STR 2016 2017 PDFDocument48 pagesPolycopie Cours Bioch STR 2016 2017 PDFAmanda LaleyePas encore d'évaluation
- LipidesDocument74 pagesLipidesالمستعينة باللهPas encore d'évaluation
- 1.métabolisme Des Glucides GlycolyseDocument39 pages1.métabolisme Des Glucides GlycolyseboubouothnielPas encore d'évaluation
- Cours 3 Biochimie MicrobienneDocument29 pagesCours 3 Biochimie MicrobienneSoumia BenPas encore d'évaluation
- Metabolisme Du GlycogeneDocument4 pagesMetabolisme Du GlycogeneJérémie barakaPas encore d'évaluation
- 1-Troubles Du Métabolisme Des Glucides PDFDocument5 pages1-Troubles Du Métabolisme Des Glucides PDFAisSa KeRrøùùm100% (1)
- QCM-EMD Botanique 2Document5 pagesQCM-EMD Botanique 2Mar OuaPas encore d'évaluation
- Chapitre 1 Pharmacologie Gle - 3Document30 pagesChapitre 1 Pharmacologie Gle - 3M.B. IsmailPas encore d'évaluation
- MICROBIOLOGIEDocument46 pagesMICROBIOLOGIEAlys ANdriaPas encore d'évaluation
- BIO 111 - CHAP - III - IV - Et - VDocument8 pagesBIO 111 - CHAP - III - IV - Et - VBerthe Aristide Ngue NkongoPas encore d'évaluation
- Biochimie Structurale Des Lipides PDFDocument37 pagesBiochimie Structurale Des Lipides PDFDr.Zakaria MFTPas encore d'évaluation
- Les-Enzymes-Plasmatiques TAM 2Document7 pagesLes-Enzymes-Plasmatiques TAM 2Snawa DangoaPas encore d'évaluation
- Feuilletage PDFDocument28 pagesFeuilletage PDFAbderrahim BoudaniPas encore d'évaluation
- Les Troubles du Rythme Cardiaque : Comprendre, Diagnostiquer et TraiterD'EverandLes Troubles du Rythme Cardiaque : Comprendre, Diagnostiquer et TraiterPas encore d'évaluation
- Quelle alimentation pour l'hémochromatose ?D'EverandQuelle alimentation pour l'hémochromatose ?Évaluation : 4 sur 5 étoiles4/5 (1)
- Extrait de Cours Tale SVTDocument26 pagesExtrait de Cours Tale SVTPascal GadedjissoPas encore d'évaluation
- Preuve de Mathématiques, BAC Série A4, Normale, Année 2015, TogoDocument2 pagesPreuve de Mathématiques, BAC Série A4, Normale, Année 2015, TogoPascal GadedjissoPas encore d'évaluation
- Seance 5Document17 pagesSeance 5Pascal GadedjissoPas encore d'évaluation
- 7 - Physiologie Du Muscle SquelettiqueDocument11 pages7 - Physiologie Du Muscle SquelettiquePascal GadedjissoPas encore d'évaluation
- 8 - BioénergetiqueDocument12 pages8 - BioénergetiquePascal GadedjissoPas encore d'évaluation
- Introduction " Les Antibiotiques "Document94 pagesIntroduction " Les Antibiotiques "Pascal GadedjissoPas encore d'évaluation
- Chapitre IIDocument12 pagesChapitre IIPascal GadedjissoPas encore d'évaluation
- CHM144 Chimie Organique FSS P2Document31 pagesCHM144 Chimie Organique FSS P2Pascal GadedjissoPas encore d'évaluation
- CHM144 Chimie Organique FSS P5Document76 pagesCHM144 Chimie Organique FSS P5Pascal GadedjissoPas encore d'évaluation
- COVADISDocument4 pagesCOVADISOriane RouaudPas encore d'évaluation
- SECHIMIEDocument2 pagesSECHIMIEekodolilian36Pas encore d'évaluation
- Microstrad N02Document60 pagesMicrostrad N02michel.soucillePas encore d'évaluation
- Firewalld Rule ExampleDocument1 pageFirewalld Rule ExamplefaisalPas encore d'évaluation
- J Pneumo 2009 04 002Document6 pagesJ Pneumo 2009 04 002dj1223549Pas encore d'évaluation
- Livret Merimee VenusDocument29 pagesLivret Merimee VenusanneescollegelyceePas encore d'évaluation
- Chomsky vs. Tomasello - Deux Approches Opposées de L'acquisition Du LangageDocument2 pagesChomsky vs. Tomasello - Deux Approches Opposées de L'acquisition Du LangageheliloPas encore d'évaluation
- Travaux Dirigés Ecoulement A Surface LibreDocument6 pagesTravaux Dirigés Ecoulement A Surface LibreSilué Nawa PatricePas encore d'évaluation
- L Ère Des Technocrates Le Cas Des PontsDocument2 pagesL Ère Des Technocrates Le Cas Des Pontserrab2012Pas encore d'évaluation
- Ech de Borg - Article PDFDocument18 pagesEch de Borg - Article PDFAnonymous 3bHuFMwQ4fPas encore d'évaluation
- 4 Somiem Tots Junts GENERALDocument15 pages4 Somiem Tots Junts GENERALAntoniaPas encore d'évaluation
- Article PortuguesDocument88 pagesArticle PortuguesDiana Bolanca100% (1)
- Les Experts Du Comité D'entreprise: Numéros JuridiquesDocument88 pagesLes Experts Du Comité D'entreprise: Numéros JuridiquesGhislain AmaniPas encore d'évaluation
- Droite Et Gauche - PrépaDocument7 pagesDroite Et Gauche - Prépasophie similiaPas encore d'évaluation
- SVT Dossier 3èmeDocument3 pagesSVT Dossier 3èmesohan.duongPas encore d'évaluation
- UntitledDocument262 pagesUntitledDiaba Messan100% (2)
- Chapitre 1 - Regime Neutre Et Protection Contre Les Contacts IndirectDocument13 pagesChapitre 1 - Regime Neutre Et Protection Contre Les Contacts IndirectmehPas encore d'évaluation
- Offre D'emploi-Procurement Officer PDFDocument4 pagesOffre D'emploi-Procurement Officer PDFBienvenu Best ParsyPas encore d'évaluation
- Chapitre4 LFDocument47 pagesChapitre4 LFtouzajohanPas encore d'évaluation
- Agitel LP3 Acg Impot Foncier 2019-2020Document8 pagesAgitel LP3 Acg Impot Foncier 2019-2020Dalmas DanpulloPas encore d'évaluation
- ControleAlgorithmes Guglielmi RGD 2023002Document22 pagesControleAlgorithmes Guglielmi RGD 2023002AnuasPas encore d'évaluation
- TDR Poste - Responsable Technique Du Programme de Sortie Renforcement Du Capital Humain Et Autonomisation Des Femmes VFDocument4 pagesTDR Poste - Responsable Technique Du Programme de Sortie Renforcement Du Capital Humain Et Autonomisation Des Femmes VFAbdouPas encore d'évaluation
- RECRUTEMENTS-APEV-T-.docx GRAINES DE PAIXDocument7 pagesRECRUTEMENTS-APEV-T-.docx GRAINES DE PAIXfructueuxPas encore d'évaluation
- 4 PolynomesAnnulateursDocument6 pages4 PolynomesAnnulateursChrist Rahul MouayaPas encore d'évaluation
- Etude TechniqueDocument16 pagesEtude TechniqueMarsoPas encore d'évaluation
- Pfi NégoDocument6 pagesPfi Négoraphael melochePas encore d'évaluation
- Constitution Du MoteurDocument21 pagesConstitution Du MoteurAbdo Abdo100% (1)
- L3 - Economie Du Travail S1 N 02Document33 pagesL3 - Economie Du Travail S1 N 02paulmariedufourPas encore d'évaluation
- Approche SystemiqueDocument5 pagesApproche SystemiqueNïna MrzkPas encore d'évaluation