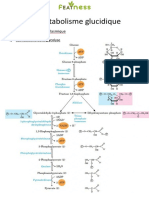
Cycle de Krebs
Cycle de Krebs
Télécharger au format pptx, pdf ou txt
Vous aimerez peut-être aussi
- UE 6.1 TD Presentation Document Le Soin Sous Tension - PDF OkDocument32 pagesUE 6.1 TD Presentation Document Le Soin Sous Tension - PDF Okks7b5xxsnhPas encore d'évaluation
- Presses Universitaires de France La Revue AdministrativeDocument9 pagesPresses Universitaires de France La Revue AdministrativeNicolas Diaz-CruzPas encore d'évaluation
- Metabolismes Des Glucides PDFDocument151 pagesMetabolismes Des Glucides PDFMa RyPas encore d'évaluation
- Poly 03-Metabolisme Des GlucidesDocument52 pagesPoly 03-Metabolisme Des Glucidessafemind100% (1)
- Pharmacocinetique QualitativeDocument45 pagesPharmacocinetique Qualitativecep100% (4)
- Biosynthése Des Lipidsynt10Document9 pagesBiosynthése Des Lipidsynt10mhichePas encore d'évaluation
- Voie Des Pentoses PhosphatesDocument26 pagesVoie Des Pentoses Phosphatescep100% (1)
- CH3.Lipogénèse PFDocument25 pagesCH3.Lipogénèse PFkhammar hichemPas encore d'évaluation
- Cycle de Krebs PDFDocument9 pagesCycle de Krebs PDFMa RyPas encore d'évaluation
- CHAP7 MET DES LIPIDES 1 Métabolisme Des Acides Gras 2022 2023 PLUSDocument9 pagesCHAP7 MET DES LIPIDES 1 Métabolisme Des Acides Gras 2022 2023 PLUSdianaPas encore d'évaluation
- Metabolisme LipidesDocument49 pagesMetabolisme LipidespartiraretirapasPas encore d'évaluation
- Cycle de KrebsDocument36 pagesCycle de Krebskeraghel nourPas encore d'évaluation
- Glucides: Les Grands Articles d'UniversalisD'EverandGlucides: Les Grands Articles d'UniversalisPas encore d'évaluation
- Métabolisme LipidiqueDocument2 pagesMétabolisme LipidiqueAro Manda100% (1)
- Le Cycle de KrebsDocument19 pagesLe Cycle de KrebsLandry RimbaybarPas encore d'évaluation
- GlycolyseDocument45 pagesGlycolysecepPas encore d'évaluation
- Metabo Partie2Document6 pagesMetabo Partie2la biochimiste100% (2)
- Métabolisme Des GlucidesDocument6 pagesMétabolisme Des GlucidesHouda SoniaPas encore d'évaluation
- Métabolisme Des Acide AminésDocument26 pagesMétabolisme Des Acide Aminés[AE]100% (4)
- 2 - Métabolisme Des Lipides PDFDocument30 pages2 - Métabolisme Des Lipides PDFFanny GrouxPas encore d'évaluation
- Métabolisme PhosphocalciqueDocument7 pagesMétabolisme Phosphocalciquekaka lotyPas encore d'évaluation
- Metabolisme Des Glucides PDFDocument157 pagesMetabolisme Des Glucides PDFMariemBoujmal33% (3)
- Physiologie de L'équilibre Acide-BaseDocument31 pagesPhysiologie de L'équilibre Acide-BaseSOLTANIPas encore d'évaluation
- Guide GlycogénoseDocument73 pagesGuide GlycogénoseVeronique HamelPas encore d'évaluation
- NeoglucogeneseDocument35 pagesNeoglucogenesecepPas encore d'évaluation
- Acides NucleiquesDocument31 pagesAcides NucleiquesLaila Home0% (1)
- Metabolisme Des TriglycéridesDocument24 pagesMetabolisme Des Triglycéridesvague2000Pas encore d'évaluation
- Catabolisme Des Lipides Complexes L3 BiochimieDocument22 pagesCatabolisme Des Lipides Complexes L3 Biochimiebiochimie L3Pas encore d'évaluation
- Métabolisme Des GlucidesDocument28 pagesMétabolisme Des GlucidesilaPas encore d'évaluation
- Métabolisme Des Glucides Fiche PDFDocument7 pagesMétabolisme Des Glucides Fiche PDFNaïs LABBADIPas encore d'évaluation
- Métabolisme Des LipoproteinesDocument13 pagesMétabolisme Des LipoproteinescepPas encore d'évaluation
- Exploration Du Métabolisme Glucidique Et Maladies DiabétiquesDocument78 pagesExploration Du Métabolisme Glucidique Et Maladies DiabétiquesFiston KabambaPas encore d'évaluation
- 3 - Metabolisme Des ProtidesDocument14 pages3 - Metabolisme Des ProtidesPascal GadedjissoPas encore d'évaluation
- Metabolisme Des Acides AminesDocument12 pagesMetabolisme Des Acides AminesWissal ElbarPas encore d'évaluation
- 12-Les CatecholaminesDocument4 pages12-Les CatecholaminesoutsiderPas encore d'évaluation
- 1 Glucides Medecine 1 1Document43 pages1 Glucides Medecine 1 1Ali SegueniPas encore d'évaluation
- METABOLISME DU GLYCOGENE - PDFXDocument56 pagesMETABOLISME DU GLYCOGENE - PDFXAmadou Marouba SowPas encore d'évaluation
- Métabolisme Des Acides AminésDocument40 pagesMétabolisme Des Acides AminéscepPas encore d'évaluation
- Bioch2an27 NeoglucogeneseDocument9 pagesBioch2an27 NeoglucogeneseIngenieur AgroPas encore d'évaluation
- Métabolisme Des Glucides DR - Khebbat Zerargui F.Document68 pagesMétabolisme Des Glucides DR - Khebbat Zerargui F.oumnia mairechePas encore d'évaluation
- Bêta OxydationDocument7 pagesBêta OxydationAnass MaaroufiPas encore d'évaluation
- La NéoglucogenèseDocument34 pagesLa Néoglucogenèsevague2000Pas encore d'évaluation
- Biochimie2an-Equilibre Acidobasique2021demmakDocument15 pagesBiochimie2an-Equilibre Acidobasique2021demmakGlobule RougePas encore d'évaluation
- 1 Acide BaseDocument4 pages1 Acide BaseSalifou Abdoul Nasser100% (1)
- Chap 1 Les Glucides Structure Et MétabolismeDocument75 pagesChap 1 Les Glucides Structure Et Métabolismeafaf gou100% (1)
- Les Acides AminesDocument7 pagesLes Acides Aminesbouguidima_sanaPas encore d'évaluation
- BiochimieDocument109 pagesBiochimieedgardokafPas encore d'évaluation
- Metabolisme Du CholesterolDocument19 pagesMetabolisme Du Cholesterol130660LYC OMAR BENABDELAZIZ NEDROMAPas encore d'évaluation
- NeoglucogeneseDocument29 pagesNeoglucogeneseHassan ElabdiPas encore d'évaluation
- 2 - Le Métabolisme Des GlucidesDocument12 pages2 - Le Métabolisme Des GlucidesFanny Groux100% (2)
- TD 2 Cycle de KrebsDocument1 pageTD 2 Cycle de KrebsDjawed Bout100% (1)
- Biochimie CliniqueDocument9 pagesBiochimie CliniqueMACON824Pas encore d'évaluation
- Métabolisme Des Acides AminésDocument87 pagesMétabolisme Des Acides AminésBlueskyindan100% (1)
- Glucides 1Document47 pagesGlucides 1Rania MaddahPas encore d'évaluation
- MEtabolisme Des GlucidesDocument16 pagesMEtabolisme Des GlucidesOtmane HajiPas encore d'évaluation
- Enzymologie 9Document31 pagesEnzymologie 9Dr.Zakaria MFTPas encore d'évaluation
- Métabolisme Du Cholestérol-Converti (1) - 2Document10 pagesMétabolisme Du Cholestérol-Converti (1) - 2Bouazzi Islemm100% (1)
- Physiologie Du Globule Rouge Et Physiopathologie Des AnémiesDocument87 pagesPhysiologie Du Globule Rouge Et Physiopathologie Des AnémiesChahine BlPas encore d'évaluation
- Les Troubles du Rythme Cardiaque : Comprendre, Diagnostiquer et TraiterD'EverandLes Troubles du Rythme Cardiaque : Comprendre, Diagnostiquer et TraiterPas encore d'évaluation
- Applications de la spectrophotomérie en phytochimie: sciencesD'EverandApplications de la spectrophotomérie en phytochimie: sciencesPas encore d'évaluation
- Pharmacologie Du Système Nerveu SympathiqueDocument88 pagesPharmacologie Du Système Nerveu SympathiquecepPas encore d'évaluation
- Suivi Thérapeutique PharmacologiqueDocument40 pagesSuivi Thérapeutique Pharmacologiquecep75% (4)
- Médicaments Et GrossesseDocument33 pagesMédicaments Et GrossessecepPas encore d'évaluation
- TD ImputabilitéDocument38 pagesTD ImputabilitécepPas encore d'évaluation
- Ordonnance MédicaleDocument52 pagesOrdonnance MédicalecepPas encore d'évaluation
- Médicaments Et AllaitementDocument30 pagesMédicaments Et AllaitementcepPas encore d'évaluation
- Recepto LogieDocument33 pagesRecepto LogiecepPas encore d'évaluation
- Physiologie DigestiveDocument26 pagesPhysiologie DigestivecepPas encore d'évaluation
- Physiologie RespiratoireDocument102 pagesPhysiologie Respiratoirecep100% (4)
- Physiologie Du Système NerveuxDocument85 pagesPhysiologie Du Système Nerveuxcep100% (1)
- Physiologie Endocrine Partie 3Document48 pagesPhysiologie Endocrine Partie 3cep100% (1)
- Conditionnement PharmaceutiqueDocument51 pagesConditionnement Pharmaceutiquecep100% (4)
- Pharmacocinetique QuantitativeDocument29 pagesPharmacocinetique Quantitativecep100% (2)
- Les Glandes EndocrinesDocument44 pagesLes Glandes EndocrinescepPas encore d'évaluation
- Physiologie Endocrine Partie 1Document36 pagesPhysiologie Endocrine Partie 1cep100% (1)
- Voie AdministrationDocument41 pagesVoie Administrationcep100% (1)
- Physio Renale 4eme CoursDocument80 pagesPhysio Renale 4eme CourscepPas encore d'évaluation
- Le Systeme de Santé MarocainDocument32 pagesLe Systeme de Santé Marocaincep86% (7)
- Biologie MoléculaireDocument291 pagesBiologie Moléculairecep100% (6)
- Métabolisme Des LipoproteinesDocument13 pagesMétabolisme Des LipoproteinescepPas encore d'évaluation
- CRMDocument40 pagesCRMcepPas encore d'évaluation
- Introduction Sur Le Métabolisme LipidesDocument21 pagesIntroduction Sur Le Métabolisme Lipidescep100% (1)
- TD2Document15 pagesTD2cepPas encore d'évaluation
- Catabolisme Acides GrasDocument11 pagesCatabolisme Acides GrascepPas encore d'évaluation
- NeoglucogeneseDocument35 pagesNeoglucogenesecepPas encore d'évaluation
- Geographie Mers Oceans PDFDocument2 pagesGeographie Mers Oceans PDFNlPas encore d'évaluation
- Searle Subjectif&Objectif PDFDocument1 pageSearle Subjectif&Objectif PDFDavid Cruz FerrerPas encore d'évaluation
- Bernard PalissyDocument15 pagesBernard PalissyNicolas BowerPas encore d'évaluation
- Transparence Des SalairesDocument2 pagesTransparence Des SalairesRaph AelPas encore d'évaluation
- 62 FRN62 0527Document24 pages62 FRN62 0527christianmakossoPas encore d'évaluation
- Exercice S Grad Div RotDocument2 pagesExercice S Grad Div RotMalak arabPas encore d'évaluation
- Trends Tendances Une Poubelle IntelligenteDocument2 pagesTrends Tendances Une Poubelle IntelligenteArmand OmatukuPas encore d'évaluation
- MICROALBUMINEDocument3 pagesMICROALBUMINECheikh Ismaïla BAPas encore d'évaluation
- A Les Fonctions VitalesDocument13 pagesA Les Fonctions VitalesDiopPas encore d'évaluation
- Nouveaux Produits Alimentaires PDFDocument35 pagesNouveaux Produits Alimentaires PDFMimoun Kandoussi100% (1)
- La Contribution Du Contrôle Budgétaire À La Performance de L'entreprise (Groupe 11)Document43 pagesLa Contribution Du Contrôle Budgétaire À La Performance de L'entreprise (Groupe 11)Omaima Es-salehyPas encore d'évaluation
- Cours de Recherche Bibliographique PDFDocument46 pagesCours de Recherche Bibliographique PDFSeif15100% (1)
- 3-6 Nerf IOA Cours DIU 2023Document18 pages3-6 Nerf IOA Cours DIU 2023Professeur Christian DumontierPas encore d'évaluation
- Doua QumaylDocument9 pagesDoua QumaylJohn Lukusa Aboubakar100% (2)
- 8 Vérités Sur Le Métabolisme Pour Brûler Des CaloriesDocument13 pages8 Vérités Sur Le Métabolisme Pour Brûler Des CaloriesPierre JehannoPas encore d'évaluation
- Markale Jean - Le Cycle Du Graal Volume 1 La Naissance Du Roi ArthurDocument424 pagesMarkale Jean - Le Cycle Du Graal Volume 1 La Naissance Du Roi ArthurGC Record100% (4)
- 2019-02-04-LIVRES HEBDO SUPPLEMENT-1er Fevrier 2019-10000000056033108Document2 pages2019-02-04-LIVRES HEBDO SUPPLEMENT-1er Fevrier 2019-10000000056033108louis.giraudeau.proPas encore d'évaluation
- GipDocument15 pagesGipNormandieActuPas encore d'évaluation
- Sémiologie Des DéshydratationsDocument23 pagesSémiologie Des DéshydratationsSasy SasyPas encore d'évaluation
- Tableau CIA 03 Pour Gare de Cahors ComplétéDocument3 pagesTableau CIA 03 Pour Gare de Cahors ComplétéAbigaelle DinetPas encore d'évaluation
- Cours 1. Recherche DocumenaireDocument4 pagesCours 1. Recherche DocumenairerabihaPas encore d'évaluation
- Regression MultipleDocument19 pagesRegression MultipleJawad EnnasihiPas encore d'évaluation
- CorrelationDocument9 pagesCorrelationIbrahim DadaPas encore d'évaluation
- Résumé de Histoire Des Idées Et Des Arts 17ème Siècle S2Document8 pagesRésumé de Histoire Des Idées Et Des Arts 17ème Siècle S2anas75% (8)
- Restaurer La Purete Sexuelle Dans Votre MinistereDocument3 pagesRestaurer La Purete Sexuelle Dans Votre MinistereAvengPas encore d'évaluation
- Cohesion Des Groupes SportifsDocument38 pagesCohesion Des Groupes SportifsAbderrahim Out100% (1)
- Isabelle Mellot, La Violence Du Rire Dans Le Conte Cruel Fin-De-Siècle, Chez Mirbeau Et Villiers de l'Isle-AdamDocument13 pagesIsabelle Mellot, La Violence Du Rire Dans Le Conte Cruel Fin-De-Siècle, Chez Mirbeau Et Villiers de l'Isle-AdamAnonymous 5r2Qv8aonfPas encore d'évaluation
- Marie-Helene MathieuDocument4 pagesMarie-Helene Mathieuapi-248683620Pas encore d'évaluation