Academia.edu no longer supports Internet Explorer.
To browse Academia.edu and the wider internet faster and more securely, please take a few seconds to upgrade your browser.
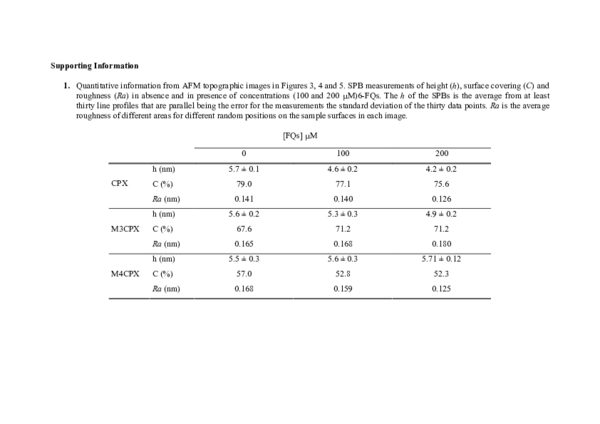
Interfacial Membrane Effects of Fluoroquinolones as Revealed by a Combination of Fluorescence Binding Experiments and Atomic Force Microscopy Observations
Interfacial Membrane Effects of Fluoroquinolones as Revealed by a Combination of Fluorescence Binding Experiments and Atomic Force Microscopy Observations
 Teresa Vinuesa
Teresa Vinuesa2006, Langmuir
Related Papers
Biochimica Et Biophysica Acta-biomembranes
Channel-closing activity of porins from Escherichia coli in bilayer lipid membranes1986 •
The opening and closing of the ompF porin from Escherichia coli JF 701 was investigated by reconstituting the purified protein into planar bilayer membranes. The electrical conductance changes across the membranes t constant potential were used to analyze the size and aggregate nature of the porin channel complexes and the relative number of opening and closing events. We found that, when measured at pH 5.5, the channel conductance diminished and the number of closing events increased when the voltage was greater than 100 mV. The results suggest that the number of sized conductance channesls increases above this potential. There was also an increase in the smaller subunits and in the closing events when the pH was lowered to 3.5, and these changes were further enhanced by increasing the voltage. We propose that both lowering the pH and elevating the potential across the membrane stabilize the porin in a conformation n which the subunits are less tightly associated and the subunits open in a non-cooperative manner. These same conditions also appear to stabilize the closed state of the pore.
Biochimica et Biophysica Acta (BBA) - Biomembranes
Formation of large, ion-permeable membrane channels by the matrix protein (porin) of Escherichia coli1978 •
Annals of the New York Academy of Sciences
Pore Formation by the Matrix Protein (Porin) of Escherichia Coli in Planar Bilayer Membranes1980 •
European Journal of Biochemistry
The voltage-dependent activity of Escherichia coli porins in different planar bilayer reconstitutions1989 •
Journal of Bacteriology
Ion selectivity of gram-negative bacterial porins1985 •
Twelve different porins from the gram-negative bacteria Escherichia coli, Salmonella typhimurium, Pseudomonas aeruginosa, and Yersinia pestis were reconstituted into lipid bilayer membranes. Most of the porins, except outer membrane protein P, formed large, water-filled, ion-permeable channels with a single-channel conductance between 1.5 and 6 nS in 1 M KCl. The ions used for probing the pore structure had the same relative mobilities while moving through the porin pore as they did while moving in free solution. Thus the single-channel conductances of the individual porins could be used to estimate the effective channel diameters of these porins, yielding values ranging from 1.0 to 2.0 nm. Zero-current potential measurements in the presence of salt gradients across lipid bilayer membranes containing individual porins gave results that were consistent with the conclusions drawn from the single-channel experiments. For all porins except protein P, the channels exhibited a greater cat...
Biochimica et Biophysica Acta (BBA) - Biomembranes
Properties of the porin of Haemophilus influenzae type b in planar lipid bilayer membranes1986 •
The major outer membrane protein (40 kDa) of the bacterium Haemophilus influenzae type b is a porin which forms transmembrane permeability channels. It has an exclusion limit for oligosaccharides of about 1.4 kDa. When this protein was added to the aqueous phase which was bathing a planar lipid bilayer, it caused the conductance of the membrane to increase by several orders of magnitude. At low protein concentrations (2-10 pM), the conductance of the membrane increased in a stepwise fashion with an average single-channel conductance of 1.1 nS in 1 M KCl. Single-channel experiments were performed with a variety of different salts. The conductance of single channels was proportional to the specific conductance of the aqueous solution which was bathing the membrane. Current through the pores was proportional to the applied voltage, indicating that these pores are not voltage-controlled. The 40 kDa porin was very slightly cation-selective: the pores were about 1.6-times more permeable to potassium ions than to chloride ions. These properties of the 40 kDa porin are those of large water-filled channels and are characteristic of most bacterial porins. The single-channel conductance of the porin is, however, much smaller than might be expected from its exclusion limit. A model is proposed which could explain the differences in apparent pore size.
Biochimica Et Biophysica Acta - Biomembranes
Ionic selectivity of pores formed by the matrix protein (porin) of Escherichia coli1979 •
Journal of Bacteriology
Pore formation by LamB of Escherichia coli in lipid bilayer membranes1986 •
Lipid bilayer experiments were performed in the presence of different Escherichia coli LamB preparations. These LamB preparations formed two types of pores in the membranes. Large pores, which had a single-channel conductance of 2.7 nS and comprised about 1 to 6% of the total pores, were presumably contaminants which might have been induced together with LamB. LamB itself formed small pores with a single-channel conductance of 160 pS in 1 M KCl. These pores could be completely blocked by the addition of maltose and maltodextrins. Titration of the pore conductance with maltotriose suggested that there was a binding site inside the pores with a Ks of 2.5 X 10(-4) M for maltotriose. On the basis of our data we concluded that the structure of the LamB channels is quite different from the structures of the channels of general diffusion porins, such as OmpF and OmpC.
RELATED PAPERS
2024 •
Analyses de l'IRFAM
Risques psychologiques subis par les migrantes avant, pendant et après le parcours migratoire2022 •
Πρακτικά Selected Papers, ISBN 978-960-98062-8-2
H Ελληνική στην Κωνσταντινούπολη: επαπειλούμενη γλώσσα;2020 •
Behavioral and Brain Sciences
Concrete magnitudes: From numbers to time2009 •
2007 •
JIPI (Jurnal Ilmu Perpustakaan dan Informasi)
Survei Kepuasan Pengguna terhadap Kualitas Layanan Sirkulasi di Perpustakaan Umum Daerah Provinsi DKI Jakarta2023 •
2020 •
Bulletin of Engineering Geology and the Environment
Historical earthquakes that damaged Hierapolis and Laodikeia antique cities and their implications for earthquake potential of Denizli basin in western Turkey2015 •
International Journal on Semantic Web and Information Systems
SPedia2017 •
Dhaka University Journal of Pharmaceutical Sciences
Physicochemical Characterization of Artemether-Entrapped Solid Lipid Microparticles Prepared from Templated- Compritol and Capra hircus (Goat Fat) Homolipid2021 •
RELATED TOPICS
- Find new research papers in:
- Physics
- Chemistry
- Biology
- Health Sciences
- Ecology
- Earth Sciences
- Cognitive Science
- Mathematics
- Computer Science