
Tissue-Specific Tolerance to High-Temperature and Nutrient-Poor Conditions in a Canopy-Forming Macroalga, Surviving at an Ocean Warming Hotspot
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Seasonal Changes in Seawater Temperature and Shoot Length
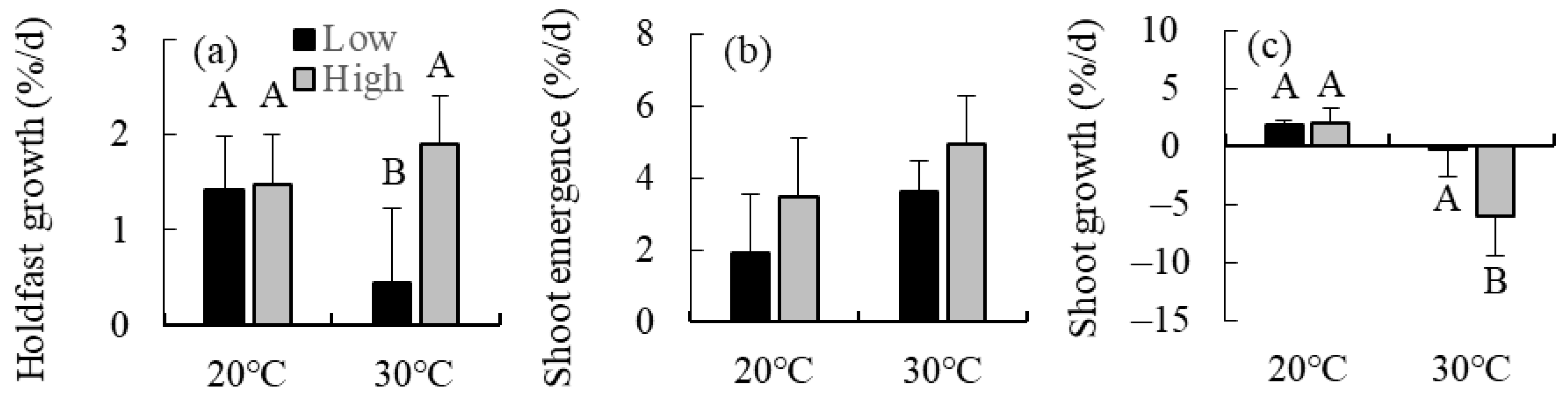
2.2. Combined Effects of Temperature and Irradiance
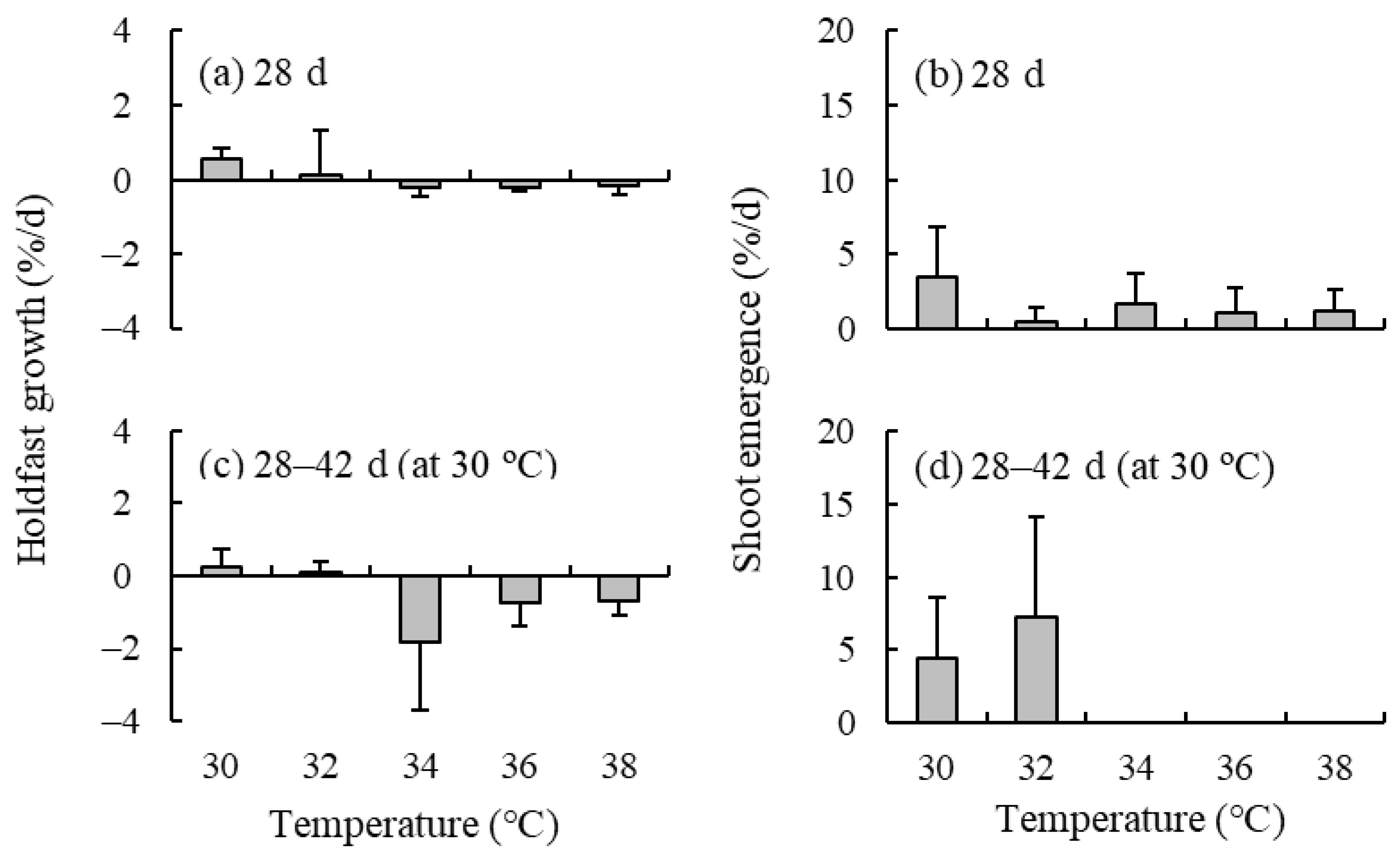
2.3. Effect of High Temperature
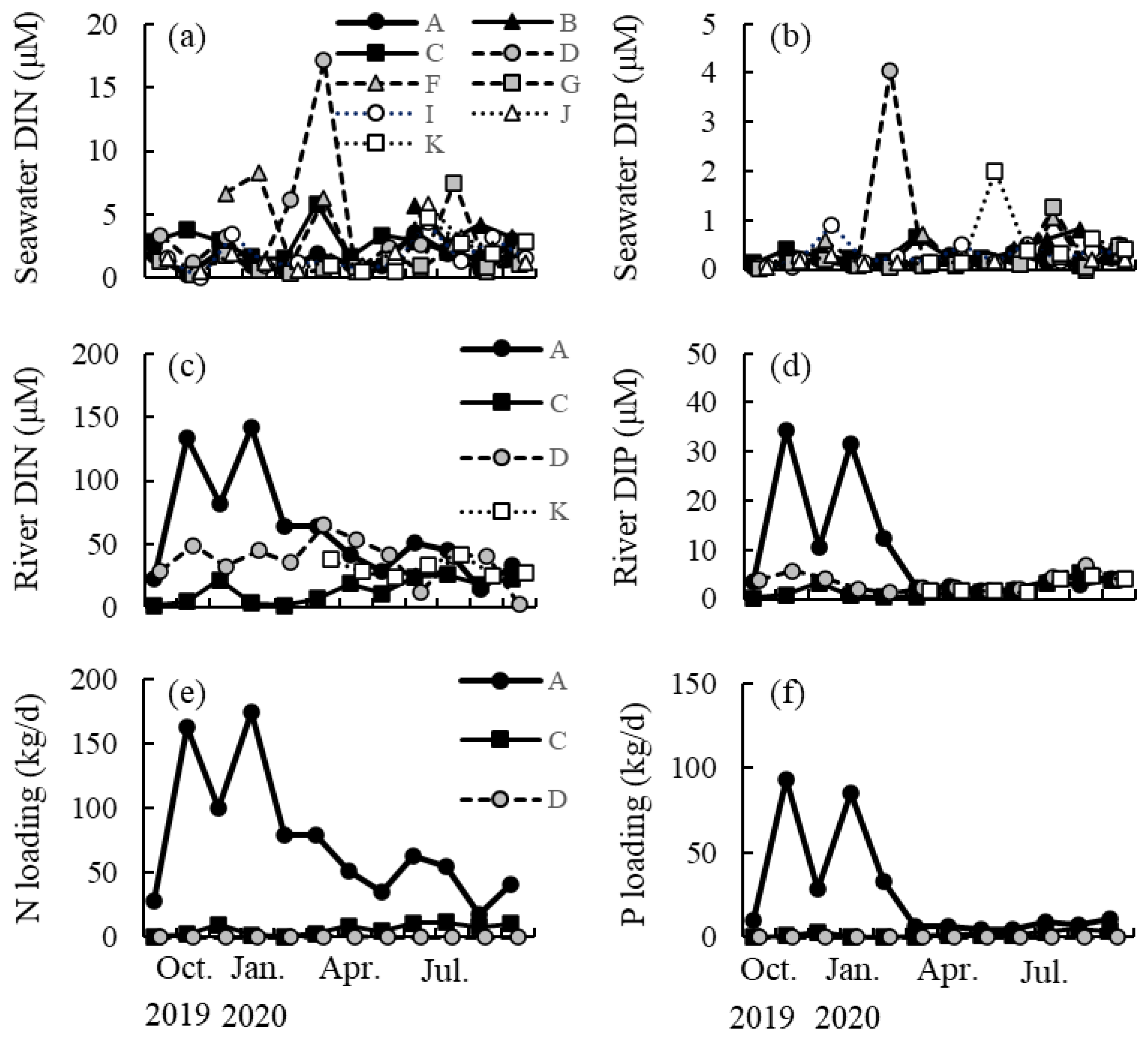
2.4. Seasonal Changes in DIN and DIP Concentrations
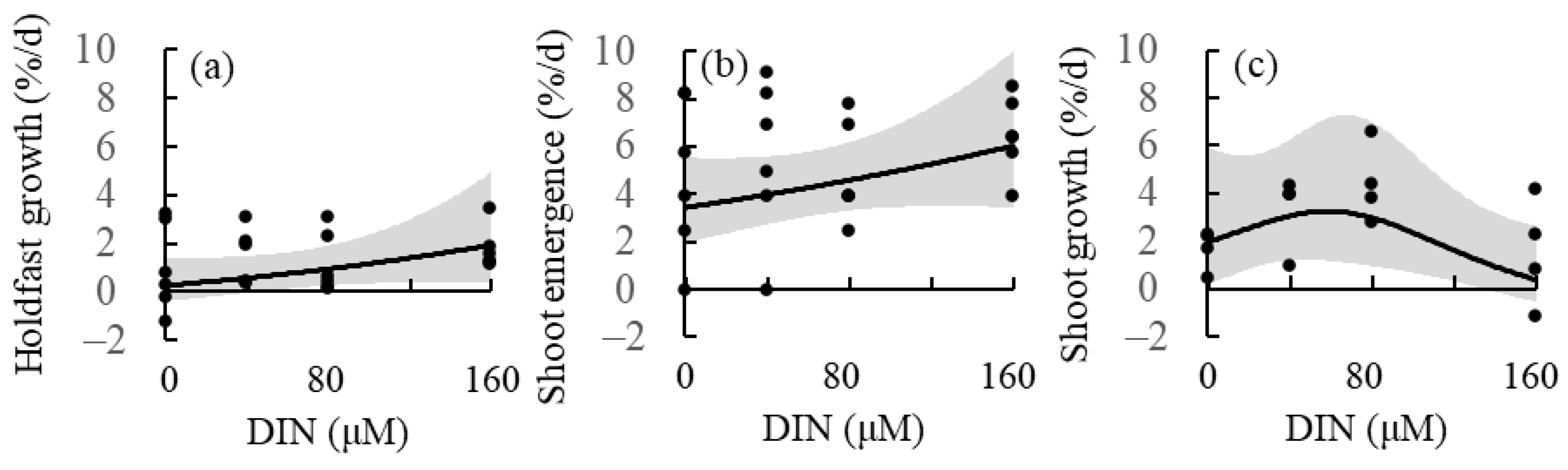
2.5. Effect of Nutrient Concentration
3. Discussion
4. Materials and Methods
4.1. Seasonal Changes in Seawater Temperature and Shoot Length
4.2. Combined Effects of Temperature and Irradiance
4.3. Effect of High Temperature
4.4. Seasonal Changes in DIN and DIP Concentrations
4.5. Effects of Nutrient Concentration
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Steneck, R.S.; Graham, M.H.; Bourque, B.J.; Corbett, D.; Erlandson, J.M.; Estes, J.A.; Tegner, M.J. Kelp forest ecosystems: Biodiversity, stability, resilience and future. Environ. Conserv. 2002, 29, 436–459. [Google Scholar] [CrossRef]
- Coleman, M.A.; Wernberg, T. Forgotten underwater forests: The key role of fucoids on Australian temperate reefs. Ecol. Evol. 2017, 7, 8406–8418. [Google Scholar] [CrossRef] [PubMed]
- Wernberg, T.; Filbee-Dexter, K. Missing the marine forest for the trees. Mar. Ecol. Prog. Ser. 2019, 612, 209–215. [Google Scholar] [CrossRef]
- Eger, A.M.; Marzinelli, E.M.; Beas-Luna, R.; Blain, C.O.; Blamey, L.K.; Byrnes, J.E.; Carnell, P.E.; Choi, C.G.; Hessing-Lewis, M.; Kim, K.Y.; et al. The value of ecosystem services in global marine kelp forests. Nat. Commun. 2023, 14, 1894. [Google Scholar] [CrossRef] [PubMed]
- Chapman, A.R.O.; Craigie, J.S. Seasonal growth in Laminaria longicruris: Relations with dissolved inorganic nutrients and internal reserves of nitrogen. Mar. Biol. 1977, 40, 197–205. [Google Scholar] [CrossRef]
- Chapman, A.R.O.; Lindley, J.E. Seasonal growth of Laminaria solidungula in the Canadian High Arctic in relation to irradiance and dissolved nutrient concentrations. Mar. Biol. 1980, 57, 1–5. [Google Scholar] [CrossRef]
- Bolton, J.J.; Lüning, K. Optimal growth and maximal survival temperatures of Atlantic Laminaria species (Phaeophyta) in culture. Mar. Biol. 1982, 66, 89–94. [Google Scholar] [CrossRef]
- tom Dieck, I. Temperature tolerance and survival in darkness of kelp gametophytes (Laminariales, Phaeophyta): Ecological and biogeographical implications. Mar. Ecol. Prog. Ser. 1993, 100, 253. [Google Scholar] [CrossRef]
- Endo, H.; Nishigaki, T.; Yamamoto, K.; Takeno, K. Age-and size-based morphological comparison between the brown alga Sargassum macrocarpum (Heterokonta; Fucales) from different depths at an exposed coast in northern Kyoto, Japan. J. Appl. Phycol. 2013, 25, 1815–1822. [Google Scholar] [CrossRef]
- Yoshida, G.; Shimabukuro, H. Seasonal population dynamics of Sargassum fusiforme (Fucales, Phaeophyta), Suo-Oshima Is., Seto Inland Sea, Japan—Development processes of a stand characterized by high density and productivity. J. Appl. Phycol. 2017, 29, 639–648. [Google Scholar] [CrossRef]
- Endo, H.; Sugie, T.; Yonemori, Y.; Nishikido, Y.; Moriyama, H.; Ito, R.; Okunishi, S. Vegetative reproduction is more advantageous than sexual reproduction in a canopy-forming clonal macroalga under ocean warming accompanied by oligotrophication and intensive herbivory. Plants 2021, 10, 1522. [Google Scholar] [CrossRef]
- Dayton, P.K.; Tegner, M.J. Catastrophic storms, El Niño, and patch stability in a southern California kelp community. Science 1984, 224, 283–285. [Google Scholar] [CrossRef]
- Tegner, M.J.; Dayton, P.K.; Edwards, P.B.; Riser, K.L. Large-scale, low-frequency oceanographic effects on kelp forest succession: A tale of two cohorts. Mar. Ecol. Prog. Ser. 1997, 146, 117–134. [Google Scholar] [CrossRef]
- Gerard, V.A. The role of nitrogen nutrition in high-temperature tolerance of the kelp, Laminaria saccharina (Chromophyta). J. Phycol. 1997, 33, 800–810. [Google Scholar] [CrossRef]
- Gao, X.; Endo, H.; Taniguchi, K.; Agatsuma, Y. Combined effects of seawater temperature and nutrient condition on growth and survival of juvenile sporophytes of the kelp Undaria pinnatifida (Laminariales; Phaeophyta) cultivated in northern Honshu, Japan. J. Appl. Phycol. 2013, 25, 269–275. [Google Scholar] [CrossRef]
- Gao, X.; Endo, H.; Nagaki, M.; Agatsuma, Y. Growth and survival of juvenile sporophytes of the kelp Ecklonia cava in response to different nitrogen and temperature regimes. Fish. Sci. 2016, 82, 623–629. [Google Scholar] [CrossRef]
- Gao, X.; Endo, H.; Nagaki, M.; Agatsuma, Y. Interactive effects of nutrient availability and temperature on growth and survival of different size classes of Saccharina japonica (Laminariales, Phaeophyceae). Phycologia 2017, 56, 253–260. [Google Scholar] [CrossRef]
- Smale, D.A. Impacts of ocean warming on kelp forest ecosystems. New Phytol. 2020, 225, 1447–1454. [Google Scholar] [CrossRef] [PubMed]
- Vergés, A.; Steinberg, P.D.; Hay, M.E.; Poore, A.G.; Campbell, A.H.; Ballesteros, E.; Heck, K.L.; Booth, D.J.; Coleman, M.A.; Feary, D.A.; et al. The tropicalization of temperate marine ecosystems: Climate-mediated changes in herbivory and community phase shifts. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140846. [Google Scholar] [CrossRef]
- Adam, T.C.; Burkepile, D.E.; Holbrook, S.J.; Carpenter, R.C.; Claudet, J.; Loiseau, C.; Thiault, L.; Brooks, A.J.; Washburn, L.; Schmitt, R.J. Landscape-scale patterns of nutrient enrichment in a coral reef ecosystem: Implications for coral to algae phase shifts. Ecol. Appl. 2021, 31, e2227. [Google Scholar] [CrossRef]
- Kumagai, N.H.; Molinos, J.G.; Yamano, H.; Takao, S.; Fujii, M.; Yamanaka, Y. Ocean currents and herbivory drive macroalgae-to coral community shift under climate warming. Proc. Natl. Acad. Sci. USA 2018, 115, 8990–8995. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T. Marine Algae of Japan; Uchida Roukakuho: Tokyo, Japan, 1998. [Google Scholar]
- Takase, T.; Tanaka, Y. Growth, maturation of Sargassum nipponicum and oceanographic conditions on the coast of Sokodo in Hachijo-jima Island, central Japan. Aquacult. Sci. 2008, 56, 369–374. [Google Scholar]
- Baba, M. Effects of temperature and irradiance on germling growth in eight Sargassaceous species. Rep. Mar. Ecol. Res. Inst. 2007, 10, 9–20. [Google Scholar]
- Murase, N.; Abe, M.; Noda, M.; Sugiura, Y. The optimal and upper critical temperatures of Sargassum fusiforme from Yamaguchi Prefecture. J. Nat. Fish. Univ. 2015, 63, 238–243. [Google Scholar]
- Abe, H.; Sanbu, M.; Kanai, M.; Yamano, H. Gaps in knowledge about distribution, utilization, and management of Sargassaceae (Fucales, Phaeophyta) in Okinawa Island, southern Japan. Reg. Stud. Mar. Sci. 2022, 53, 102431. [Google Scholar] [CrossRef]
- Charan, H.; Inomata, E.; Endo, H.; Sato, Y.; Okumura, Y.; Aoki, M.N. Decreased irradiance and nutrient enrichment mitigate the negative effect of ocean warming on growth and biochemical compositions of a canopy-forming marine macroalga. J. Mar. Sci. Eng. 2022, 10, 479. [Google Scholar] [CrossRef]
- Noro, T.; Alcantara, L.B.; Masuda, Y. Research project on fishery management of the subtropical abalone, Haliotis diversicolor in Kagoshima University. Mem. Fac. Fish. Kagoshima Univ. 2004, 53, 37–40. [Google Scholar]
- Balfagón, D.; Sengupta, S.; Gómez-Cadenas, A.; Fritschi, F.B.; Azad, R.K.; Mittler, R.; Zandalinas, S.I. Jasmonic acid is required for plant acclimation to a combination of high light and heat stress. Plant Physiol. 2019, 181, 1668–1682. [Google Scholar] [CrossRef]
- Tait, L.W.; Schiel, D.R. Impacts of temperature on primary productivity and respiration in naturally structured macroalgal assemblages. PLoS ONE 2013, 8, e74413. [Google Scholar] [CrossRef]
- Tanaka, T.; Yoshimitsu, S.; Imayoshi, Y.; Ishige, Y.; Terada, R. Distribution and characteristics of seaweed/seagrass community in Kagoshima Bay, Kagoshima Prefecture, Japan. Nippon Suisan Gakk. 2013, 79, 20–30. [Google Scholar] [CrossRef]
- Haraguchi, H.; Murase, N.; Mizukami, Y.; Noda, M.; Yoshida, G.; Terawaki, T. The optimal and maximum temperatures of nine species of the Sargassaceae in the coastal waters of Yamaguchi Prefecture. Japan. Jpn. J. Phycol. 2005, 53, 7–13. [Google Scholar]
- Schaffelke, B.; Klumpp, D.W. Short-term nutrient pulses enhance growth and photosynthesis of the coral reef macroalga Sargassum baccularia. Mar. Ecol. Prog. Ser. 1998, 170, 95–105. [Google Scholar] [CrossRef]
- Shimabukuro, H.; Terada, R.; Sotobayashi, J.; Nishihara, G.N.; Noro, T. Phenology of Sargassum duplicatum (Fucales, Phaeophyceae) from the southern coast of Satsuma Peninsula, Kagoshima, Japan. Nippon Suisan Gakk. 2007, 73, 454–460. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Sakaguchi, Y.; Terada, R. Phenology and environmental characteristics of four Sargassum species (Fucales): S. piluliferum, S. patens, S. crispifolium, and S. alternato-pinnatum from Sakurajima, Kagoshima Bay, southern Japan. Jpn. J. Phycol. 2011, 59, 1–8. [Google Scholar]
- Nakashima, H.; Tanaka, T.; Yoshimitsu, S.; Terada, R. Phenology of three species of Sargassum (Fucales) and the long-term change of seaweed community structure from Kasasa, Kagoshima Prefecture, Japan. Jpn. J. Phycol. 2013, 61, 97–105. [Google Scholar]
- Kiriyama, T.; Noda, M.; Fujii, A. Grazing and bite marks on Eclonia kurome caused by several herbivorous fishes. Acuacult. Sci. 2001, 49, 431–438. [Google Scholar]
- Endo, H.; Fukuda, H.; Takahashi, D.; Okumura, Y.; Inomata, E.; Ito, K.; Yoshimizu, C.; Tayasu, I.; Nagata, T. Influence of isotope fractionation on the nitrogen isotope composition of the brown macroalga Undaria pinnatifida. Phycol. Res. 2018, 66, 262–268. [Google Scholar] [CrossRef]
- Endo, H.; Moriyama, H.; Okumura, Y. Photoinhibition and photoprotective responses of a brown marine macroalga acclimated to different light and nutrient regimes. Antioxidants 2023, 12, 357. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Factor | df | SS | MS | F | p | |
|---|---|---|---|---|---|---|---|
| Holdfast growth rate | T | 1 | 0.487 | 0.487 | 1.345 | 0.260 | |
| I | 1 | 3.399 | 3.399 | 9.387 | 0.006 | * | |
| T * I | 1 | 2.977 | 2.977 | 8.223 | 0.010 | * | |
| Shoot emergence rate | T | 1 | 15.39 | 15.386 | 7.771 | 0.011 | * |
| I | 1 | 12.34 | 12.342 | 6.234 | 0.021 | * | |
| T * I | 1 | 0.090 | 0.088 | 0.045 | 0.835 | ||
| Shoot growth rate | T | 1 | 104.47 | 104.470 | 22.847 | <0.001 | * |
| I | 1 | 30.040 | 30.040 | 6.570 | 0.025 | * | |
| T * I | 1 | 35.560 | 35.560 | 7.776 | 0.016 | * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Endo, H.; Kodama, M.; Kawashima, R.; Kumagai, M.; Matsuoka, M.; Ebata, K.; Okunishi, S. Tissue-Specific Tolerance to High-Temperature and Nutrient-Poor Conditions in a Canopy-Forming Macroalga, Surviving at an Ocean Warming Hotspot. Plants 2024, 13, 1689. https://doi.org/10.3390/plants13121689
Endo H, Kodama M, Kawashima R, Kumagai M, Matsuoka M, Ebata K, Okunishi S. Tissue-Specific Tolerance to High-Temperature and Nutrient-Poor Conditions in a Canopy-Forming Macroalga, Surviving at an Ocean Warming Hotspot. Plants. 2024; 13(12):1689. https://doi.org/10.3390/plants13121689
Chicago/Turabian StyleEndo, Hikaru, Masafumi Kodama, Ryoya Kawashima, Momochika Kumagai, Midori Matsuoka, Keigo Ebata, and Suguru Okunishi. 2024. "Tissue-Specific Tolerance to High-Temperature and Nutrient-Poor Conditions in a Canopy-Forming Macroalga, Surviving at an Ocean Warming Hotspot" Plants 13, no. 12: 1689. https://doi.org/10.3390/plants13121689