Monte
Monte
Download as pdf or txt
You might also like
- Audio System (Separate Type Amplifier)Document29 pagesAudio System (Separate Type Amplifier)Khanh ĐinhNo ratings yet
- AACR 2022 Proceedings: Part A Online-Only and April 10From EverandAACR 2022 Proceedings: Part A Online-Only and April 10No ratings yet
- ResilienceDocument40 pagesResilienceakhilNo ratings yet
- Prognostic Relationship of Metabolic Profile Obtained of Melanoma B16F10Document11 pagesPrognostic Relationship of Metabolic Profile Obtained of Melanoma B16F10Alvaro GaldosNo ratings yet
- 004 EAd 01Document5 pages004 EAd 01dropdeadbeautifullNo ratings yet
- Citosol (Thiamylal Sodium) Triggers Apoptosis and Affects Gene Expressions of Murine Leukemia RAW 264.7 Cells - RS-C Wu, C-S YuDocument1 pageCitosol (Thiamylal Sodium) Triggers Apoptosis and Affects Gene Expressions of Murine Leukemia RAW 264.7 Cells - RS-C Wu, C-S YuAlondra MaldonadoNo ratings yet
- The Tyrosine Kinase Inhibitor Nilotinib Is More Efficient Than Mitotane in Decreasing Cell Viability in Spheroids Prepared From Adrenocortical Carcinoma CellsDocument16 pagesThe Tyrosine Kinase Inhibitor Nilotinib Is More Efficient Than Mitotane in Decreasing Cell Viability in Spheroids Prepared From Adrenocortical Carcinoma CellsElaine SilveiraNo ratings yet
- JNCI J Natl Cancer Inst 1997 Ahmad 1881 6Document6 pagesJNCI J Natl Cancer Inst 1997 Ahmad 1881 6Satria Bayu PratamaNo ratings yet
- DOI: 10.30644/rik.v12i2.826Document5 pagesDOI: 10.30644/rik.v12i2.826janice ferdinandNo ratings yet
- Ganoderic Acid T From Ganoderma Mycelia Induces Mitochondria Mediated Apoptosis in Lung Cancer CellsDocument7 pagesGanoderic Acid T From Ganoderma Mycelia Induces Mitochondria Mediated Apoptosis in Lung Cancer CellsGandalfNo ratings yet
- Chitosan Exerts Anticancer Activity Through Induction of Apoptosis and Cell Cycle Arrest in Oral Cancer CellsDocument8 pagesChitosan Exerts Anticancer Activity Through Induction of Apoptosis and Cell Cycle Arrest in Oral Cancer CellshameedhaaNo ratings yet
- FolicacidDocument13 pagesFolicacida.mahdieh90No ratings yet
- Arango-Varela Et Al., 2021Document16 pagesArango-Varela Et Al., 2021Maria Elena MaldonadoNo ratings yet
- Journal Pre-Proof: LWT - Food Science and TechnologyDocument46 pagesJournal Pre-Proof: LWT - Food Science and TechnologyJulio MoralesNo ratings yet
- Novel Boswellic Acids Nanoparticles Induces Cell Death in Prostate Cancer CellsDocument9 pagesNovel Boswellic Acids Nanoparticles Induces Cell Death in Prostate Cancer CellsSaji UthamanNo ratings yet
- 5.13695 Biogeckoajournalfornewzealandherpetology 02-01-47Document7 pages5.13695 Biogeckoajournalfornewzealandherpetology 02-01-47mehulshah1126No ratings yet
- Bakuchiol Inhibits Cell Proliferation and Induces Apoptosis and Cell Cycle Arrest in SGC-7901 Human Gastric Cancer CellsDocument6 pagesBakuchiol Inhibits Cell Proliferation and Induces Apoptosis and Cell Cycle Arrest in SGC-7901 Human Gastric Cancer CellsFarhana AnuarNo ratings yet
- Gene Expression and Histopathology Alterations During RatDocument17 pagesGene Expression and Histopathology Alterations During RatTahir AliNo ratings yet
- 437876.int Canc Congr PaperDocument13 pages437876.int Canc Congr PaperRiani Dwi HastutiNo ratings yet
- Norazalina Et AlDocument8 pagesNorazalina Et AlHajah Sofyamarwa RachmawatiNo ratings yet
- Plant Extract - Colitis Induced Hepatic InjuryDocument9 pagesPlant Extract - Colitis Induced Hepatic InjurySuprada NatarajuNo ratings yet
- Vitamin C AutophagyDocument12 pagesVitamin C AutophagyureehwsenqNo ratings yet
- Or 26 1 185 PDFDocument7 pagesOr 26 1 185 PDFRoland_IINo ratings yet
- Food and Chemical Toxicology: Kyoung Jin Nho, Jin Mi Chun, Ho Kyoung KimDocument10 pagesFood and Chemical Toxicology: Kyoung Jin Nho, Jin Mi Chun, Ho Kyoung KimMd Jahidul IslamNo ratings yet
- PHD Thesis MetabolomicsDocument5 pagesPHD Thesis Metabolomicsf1t1febysil2100% (1)
- Wang Et Al-2012-Journal of Pineal ResearchDocument8 pagesWang Et Al-2012-Journal of Pineal ResearchDionisio MesyeNo ratings yet
- Ethanol Extract of Dunaliella Salina Induces Cell Cycle Arrest and Apoptosis in A549 Human Non-Small Cell Lung Cancer CellsDocument10 pagesEthanol Extract of Dunaliella Salina Induces Cell Cycle Arrest and Apoptosis in A549 Human Non-Small Cell Lung Cancer CellsDheerdendra ChaudharyNo ratings yet
- Hydroethanolic Extract of Solanum paniculatum L. Fruits Modulates ROS and Cytokine in Human Cell LinesDocument10 pagesHydroethanolic Extract of Solanum paniculatum L. Fruits Modulates ROS and Cytokine in Human Cell Linesdra.ferrazapcrNo ratings yet
- Artículo 1Document8 pagesArtículo 1nmqkcwychfNo ratings yet
- Asadi Khiavi BioImpacts 2011 1 2Document7 pagesAsadi Khiavi BioImpacts 2011 1 2farhadNo ratings yet
- ????Document11 pages????sajabaniahmad101No ratings yet
- Euphorbia Tirucalli L. Modulates Myelopoiesis and Enhances TheDocument6 pagesEuphorbia Tirucalli L. Modulates Myelopoiesis and Enhances ThecarolineNo ratings yet
- Erc EndocrinologyDocument11 pagesErc EndocrinologyEsti Nur EkasariNo ratings yet
- Neo0906 0504Document7 pagesNeo0906 0504Maria FregniNo ratings yet
- Magnolol-Induced H460 Cells Death Via Autophagy But Not ApoptosisDocument9 pagesMagnolol-Induced H460 Cells Death Via Autophagy But Not ApoptosisChivis MoralesNo ratings yet
- Ssri On CancerDocument7 pagesSsri On CancerArpit SanghaniNo ratings yet
- 0 F 3175395 Beff 2 Eb 2 C 000000Document6 pages0 F 3175395 Beff 2 Eb 2 C 000000Linh NguyễnNo ratings yet
- Antitumor Activity of Cratylia Mollis Lectin Encapsulated Into LiposomesDocument11 pagesAntitumor Activity of Cratylia Mollis Lectin Encapsulated Into LiposomesAlbertoNo ratings yet
- Structure-Activity Relationships of Synthetic Analogs of (-) - Epigallocatechin-3-Gallate As Proteasome InhibitorsDocument12 pagesStructure-Activity Relationships of Synthetic Analogs of (-) - Epigallocatechin-3-Gallate As Proteasome InhibitorsGhibli GinNo ratings yet
- Natural Bioactive From Euphorbia Umbellata Latex As A Modulator of Mitochondrial Activity in Breast Cancer CellsDocument7 pagesNatural Bioactive From Euphorbia Umbellata Latex As A Modulator of Mitochondrial Activity in Breast Cancer CellsLaertty CabralNo ratings yet
- CPG Island Hypermethylation-Associated Silencing of Micrornas Promotes Human Endometrial CancerDocument10 pagesCPG Island Hypermethylation-Associated Silencing of Micrornas Promotes Human Endometrial CancerFerdina NidyasariNo ratings yet
- Sc05a06s27Document8 pagesSc05a06s27Achille BroggiNo ratings yet
- Mehmeti Et Al, 2011Document8 pagesMehmeti Et Al, 2011olindacabralNo ratings yet
- Efectul de Prelucrare C-Âldur-Â Pe Detectarea PCR A Modificate Genetic (MG), Soia + N Produsele Din CarneDocument2 pagesEfectul de Prelucrare C-Âldur-Â Pe Detectarea PCR A Modificate Genetic (MG), Soia + N Produsele Din CarneStefan AdrianNo ratings yet
- Toxins in SpicesDocument5 pagesToxins in SpicestatarasanurazvanNo ratings yet
- abstract-c.elegansa-setDocument8 pagesabstract-c.elegansa-setlucasteofarmNo ratings yet
- SB Dna RepairDocument6 pagesSB Dna RepairSean BarnesNo ratings yet
- Articol DecelularizareDocument12 pagesArticol DecelularizareOana MariaNo ratings yet
- DownloadDocument7 pagesDownloadphelomarkmagantangNo ratings yet
- Cristinalopesdocarmo 2020Document7 pagesCristinalopesdocarmo 2020Leandro DouglasNo ratings yet
- WAT Plasticity in Response To Beta3-ARDocument9 pagesWAT Plasticity in Response To Beta3-ARassaf.rudichNo ratings yet
- B-Sitosterol Activates Fas Signaling in Human Breast Cancer CellsDocument8 pagesB-Sitosterol Activates Fas Signaling in Human Breast Cancer CellsFrance Louie JutizNo ratings yet
- Research Article Viola Tricolor Induces Apoptosis in Cancer Cells and ExhibitsDocument9 pagesResearch Article Viola Tricolor Induces Apoptosis in Cancer Cells and ExhibitsEliasDraNo ratings yet
- Ghanemi 2017Document10 pagesGhanemi 2017YESSELYS SILVERANo ratings yet
- Present As IDocument9 pagesPresent As Iaini rahmawatiNo ratings yet
- Effects of Propolis and Phenolic Acids On Triple-NDocument14 pagesEffects of Propolis and Phenolic Acids On Triple-Nriyadh jizan ُegyptian-boys-sectionNo ratings yet
- Cancer ResearchDocument8 pagesCancer ResearchAngelNo ratings yet
- Quer Ce TinaDocument13 pagesQuer Ce Tinanorociel8132No ratings yet
- Selectivity Index of Alpinia Galanga Extract and 1'-Acetoxychavicol Acetate On Cancer Cell LinesDocument6 pagesSelectivity Index of Alpinia Galanga Extract and 1'-Acetoxychavicol Acetate On Cancer Cell LinesasrilNo ratings yet
- Thesis MetabolomicsDocument11 pagesThesis Metabolomicsfqh4d8zf100% (2)
- Stem Cells in Birth Defects Research and Developmental ToxicologyFrom EverandStem Cells in Birth Defects Research and Developmental ToxicologyTheodore P. RasmussenNo ratings yet
- Endophyte Biotechnology: Potential for Agriculture and PharmacologyFrom EverandEndophyte Biotechnology: Potential for Agriculture and PharmacologyAlexander SchoutenNo ratings yet
- Acute Limb Ischaemia: Dr. Joel Arudchelvam Consultant Vascular and Transplant Surgeon Teaching Hospital AnuradhapuraDocument30 pagesAcute Limb Ischaemia: Dr. Joel Arudchelvam Consultant Vascular and Transplant Surgeon Teaching Hospital AnuradhapuraOmomomo781No ratings yet
- Banana Leaf DressingDocument6 pagesBanana Leaf DressingOmomomo781No ratings yet
- Surgical Treatment of Anterior Urethral Stricture DiseasesDocument9 pagesSurgical Treatment of Anterior Urethral Stricture DiseasesOmomomo781No ratings yet
- Advances in Urethral Stricture Management (Version 1 Referees: 4 Approved)Document9 pagesAdvances in Urethral Stricture Management (Version 1 Referees: 4 Approved)Omomomo781No ratings yet
- Coordination of Pps Samples Over TimeDocument14 pagesCoordination of Pps Samples Over TimeLefeneNo ratings yet
- 667460-mark-scheme-pure-mathematics-and-mechanicsDocument14 pages667460-mark-scheme-pure-mathematics-and-mechanicsmuhammedzonaid304No ratings yet
- 14cyl For RTA96CDocument2 pages14cyl For RTA96Cmarko4096No ratings yet
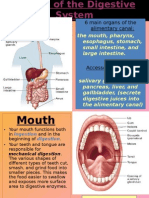
- Organs of The Digestive SystemDocument16 pagesOrgans of The Digestive SystemBrandon HunterNo ratings yet
- Everything You Need to Know About the BHMCT Programme at K.R. Mangalam University (1)Document7 pagesEverything You Need to Know About the BHMCT Programme at K.R. Mangalam University (1)K.R. Mangalam UniversityNo ratings yet
- Curriculum Vitae of Hafiz Sikander 1Document7 pagesCurriculum Vitae of Hafiz Sikander 1hafiz2422No ratings yet
- Sun 800FDocument8 pagesSun 800Fdayana paredesNo ratings yet
- 1st Lecture On The Histology of Skin by Dr. RoomiDocument14 pages1st Lecture On The Histology of Skin by Dr. RoomiMudassar RoomiNo ratings yet
- iMAC300: High-End 3-Ch ECG MachineDocument2 pagesiMAC300: High-End 3-Ch ECG MachineAsep MustopaNo ratings yet
- Common Oral Antimicrobial Therapy Dosage AdjustmentDocument2 pagesCommon Oral Antimicrobial Therapy Dosage Adjustmentnoviantyramadhani12No ratings yet
- 4 SR15 Chapter2 SM High ResDocument50 pages4 SR15 Chapter2 SM High ResMaxNo ratings yet
- 4 Complex NumDocument28 pages4 Complex NumeshaNo ratings yet
- Dying To Be Men - Psychosocial, Environmental, and Biobehavioral Directions in Promoting The Health of Men and Boys (The Routledge Series On Counseling and Psychotherapy With Boys and Men) (PDFDrive)Document527 pagesDying To Be Men - Psychosocial, Environmental, and Biobehavioral Directions in Promoting The Health of Men and Boys (The Routledge Series On Counseling and Psychotherapy With Boys and Men) (PDFDrive)kajal makwanaNo ratings yet
- Experimental and Numerical Investigations On Seismic Responses of Reinforced Concrete Structures Considering Strain Rate EffectDocument15 pagesExperimental and Numerical Investigations On Seismic Responses of Reinforced Concrete Structures Considering Strain Rate EffectIon SococolNo ratings yet
- IMS Application Form and Consent Form215 3Document3 pagesIMS Application Form and Consent Form215 3Eugen PituscanNo ratings yet
- Student Health Record: PART I: To Be Answered by Parents or GuardianDocument4 pagesStudent Health Record: PART I: To Be Answered by Parents or GuardianDerick DalisayNo ratings yet
- Barthes, Roland - MythologiesDocument183 pagesBarthes, Roland - Mythologiessaad100% (3)
- Guidelines For Using VCI MaterialDocument2 pagesGuidelines For Using VCI MaterialRajkumar ModiNo ratings yet
- cp15 - StandardDocument1 pagecp15 - Standardinfratech engineeringNo ratings yet
- What A Spa Consultant Does PDFDocument3 pagesWhat A Spa Consultant Does PDFFitzgerald PachecoNo ratings yet
- Screenshot 2023-01-31 at 10.37.57 AMDocument176 pagesScreenshot 2023-01-31 at 10.37.57 AMalhussain asiriNo ratings yet
- ISA 210 Agreeing The Terms of Audit Engagement AND LAWS AND REGULATIONSDocument4 pagesISA 210 Agreeing The Terms of Audit Engagement AND LAWS AND REGULATIONSneelumshahzaadiNo ratings yet
- My Vocabulary..... CCCDocument5 pagesMy Vocabulary..... CCCMuhammad AffanNo ratings yet
- WHQ Character - Witch Hunter (Complete, Searchable)Document25 pagesWHQ Character - Witch Hunter (Complete, Searchable)vernerNo ratings yet
- AWS-Pre-During-Post-Weld-Inspections ChecksheetDocument4 pagesAWS-Pre-During-Post-Weld-Inspections ChecksheetJohn TerryNo ratings yet
- MSBSHSE Class 9 Defence Studies TextbookDocument66 pagesMSBSHSE Class 9 Defence Studies Textbookvarshabalgude11100% (1)
- The Blood Is The Life: Haverford CollegeDocument30 pagesThe Blood Is The Life: Haverford CollegeHaneen Al IbrahimNo ratings yet
- The Mariner's Handbook NP100Document385 pagesThe Mariner's Handbook NP100mSh--No ratings yet