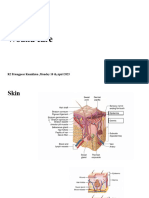
Wound healing
Wound healing
Download as pptx, pdf, or txt
You might also like
- Wound Healing SchwartzDocument37 pagesWound Healing SchwartzCarla Putri Chandra100% (1)
- Lewis Medical Surgical ch12-2Document8 pagesLewis Medical Surgical ch12-2katwoman1550% (2)
- Boik Natural Compounds in Cancer Therapy Promising Nontoxic Antitumor Agents From Plants & Other Natural Sources (2001) PDFDocument541 pagesBoik Natural Compounds in Cancer Therapy Promising Nontoxic Antitumor Agents From Plants & Other Natural Sources (2001) PDFIoana Antonesi100% (1)
- Chap-1a Wound HealingDocument60 pagesChap-1a Wound HealingAlex HaileyesusNo ratings yet
- Wound Healing, Tissue Repair, and FibrosisDocument28 pagesWound Healing, Tissue Repair, and FibrosisRibka TheodoraNo ratings yet
- Wound and Wound HealingDocument28 pagesWound and Wound HealingObehi Eromosele0% (1)
- Wound Healing and Wound CareDocument15 pagesWound Healing and Wound CareNinaNo ratings yet
- Wound Healing, Tissue Repair, and FibrosisDocument28 pagesWound Healing, Tissue Repair, and FibrosisRibka Theodora100% (1)
- Leoni 2015Document10 pagesLeoni 2015Yunita SaharawatiNo ratings yet
- Wound Healing JurnalDocument10 pagesWound Healing JurnalBrilliantNo ratings yet
- Abd 91 05 0614Document7 pagesAbd 91 05 0614Selvy Anriani GasperszNo ratings yet
- Woundcare Monday10423Document63 pagesWoundcare Monday10423Santanico De CVT deozaNo ratings yet
- Wound Healing Normal and Abnormal Mechanisms and Closure TechniquesDocument40 pagesWound Healing Normal and Abnormal Mechanisms and Closure TechniquesAhmad Fakhrozi Helmi100% (1)
- Wound Healing and Its Impairment in The Diabetic Foot: ReviewDocument9 pagesWound Healing and Its Impairment in The Diabetic Foot: ReviewJoey TsaiNo ratings yet
- 7. WoundsDocument44 pages7. Woundsapolloniwagaba02No ratings yet
- Healing and RepairDocument40 pagesHealing and RepairfromherwindowNo ratings yet
- Granulation Tissue 13102015Document33 pagesGranulation Tissue 13102015Widi Widurai100% (1)
- Wound Healing: Monisha B. Subhashree GDocument35 pagesWound Healing: Monisha B. Subhashree GGayathri MaigandanNo ratings yet
- Referat - CHRONIC WOUNDDocument19 pagesReferat - CHRONIC WOUNDAfifah Syifaul UmmahNo ratings yet
- Tissue Repair, Wound Healing and FibrosisDocument48 pagesTissue Repair, Wound Healing and FibrosisIyanAsiana100% (1)
- VfyggdbDocument33 pagesVfyggdbBibek RajNo ratings yet
- Wound Healing: Rhona Maceachen Wvsumc PgiDocument24 pagesWound Healing: Rhona Maceachen Wvsumc PgiJoher MendezNo ratings yet
- 3Wound HealingDocument35 pages3Wound HealingKhizer ShahNo ratings yet
- Chronic Inflammation Healing and RepairDocument51 pagesChronic Inflammation Healing and Repairahmed mokhtarNo ratings yet
- Basic Mollecular Wound Healing of The SkinDocument22 pagesBasic Mollecular Wound Healing of The SkindavidNo ratings yet
- Wound HealingDocument34 pagesWound HealingsadasivamagasthiyaNo ratings yet
- Wound Healing & Wound Care: Souvik Adhikari Postdoctoral TraineeDocument55 pagesWound Healing & Wound Care: Souvik Adhikari Postdoctoral TraineeClaudio Luis VenturiniNo ratings yet
- 5 - HealingDocument29 pages5 - HealingShna SaadiNo ratings yet
- Wound HealingDocument76 pagesWound Healingmannkheni209No ratings yet
- Stem CellDocument14 pagesStem CellsattwikaNo ratings yet
- Wound Healing: by Arpit VermaDocument21 pagesWound Healing: by Arpit VermaAbhishek SarafNo ratings yet
- 1 7 10 Wound HealingDocument3 pages1 7 10 Wound Healingkep1313No ratings yet
- Wounds, Wound Healing and Complications of WoundDocument36 pagesWounds, Wound Healing and Complications of WoundIbrahim AkinbolaNo ratings yet
- Wound HealingDocument46 pagesWound HealingSahin mollickNo ratings yet
- First Foundations in Pathology Part 3: Growth and RepairDocument46 pagesFirst Foundations in Pathology Part 3: Growth and RepairmadhuNo ratings yet
- Wound Repair and HealingDocument33 pagesWound Repair and Healinghetler343No ratings yet
- Wound Healing and RepairDocument54 pagesWound Healing and RepairnyangaraNo ratings yet
- Chronic InflammationDocument48 pagesChronic InflammationnravitejaNo ratings yet
- Repair & Wound Healing 2024 SMCDocument62 pagesRepair & Wound Healing 2024 SMCAhsan KazmiNo ratings yet
- Healing Gum Edited 2024Document70 pagesHealing Gum Edited 2024AMUGO JOYLYNNo ratings yet
- (SURGERY SGD) Wound HealingDocument8 pages(SURGERY SGD) Wound HealingPaulene RiveraNo ratings yet
- Tissue Repair 23Document4 pagesTissue Repair 23lidetu03No ratings yet
- Ern Mrcs Bk1Document21 pagesErn Mrcs Bk1aeages100% (1)
- Wound Healing and Perioperative Care - Vol 18 Issue 1 Feb 2006 OmfsDocument7 pagesWound Healing and Perioperative Care - Vol 18 Issue 1 Feb 2006 Omfsapi-265532519No ratings yet
- Wound Healing and Perioperative Care Vol 18 Issue 1 Feb 2006 Omfs PDFDocument7 pagesWound Healing and Perioperative Care Vol 18 Issue 1 Feb 2006 Omfs PDFR KNo ratings yet
- Regeneration and Healing 2011 DRG Neny Prasetyaningrum, MkedDocument24 pagesRegeneration and Healing 2011 DRG Neny Prasetyaningrum, MkedAnonymous BUkBUU847O100% (1)
- Wound Healing: Ziv Peled, M.DDocument8 pagesWound Healing: Ziv Peled, M.Dapi-26007957No ratings yet
- Wound Healing and Factors Affecting Wound Healing: Principles of SurgeryDocument57 pagesWound Healing and Factors Affecting Wound Healing: Principles of SurgeryAmit RamdinNo ratings yet
- Kuliah Pemulihan JaringanDocument38 pagesKuliah Pemulihan JaringanBrigitha Rahmadini PaputunganNo ratings yet
- Cells 13 00624Document13 pagesCells 13 00624bszool006No ratings yet
- 4 Chronic Inflammation IDocument38 pages4 Chronic Inflammation IAlex XanderNo ratings yet
- GROUP6Document22 pagesGROUP6AYUSHI PATELNo ratings yet
- Principles of Wound HealingDocument8 pagesPrinciples of Wound HealingTracy100% (6)
- Wound Healing and Tissue RepairDocument27 pagesWound Healing and Tissue RepairKELVIN MAYOMBONo ratings yet
- Tissue Injury and Healing: Brent Kincaid, DDS, John P. Schmitz, DDS, PHDDocument10 pagesTissue Injury and Healing: Brent Kincaid, DDS, John P. Schmitz, DDS, PHDAmith HadhimaneNo ratings yet
- Hatim NewDocument50 pagesHatim NewJaser YaminNo ratings yet
- Healing and RepairDocument50 pagesHealing and RepairVrushali BhoirNo ratings yet
- Wound HealingDocument70 pagesWound HealingSachin KpNo ratings yet
- 6.chronic Inflammation Wound Healing and Repair CMTDocument50 pages6.chronic Inflammation Wound Healing and Repair CMTemakinu19No ratings yet
- Chronic Inflammation Healing and Repair CMTDocument35 pagesChronic Inflammation Healing and Repair CMTemakinu19No ratings yet
- Complications of Third Molar Surgery JCDocument25 pagesComplications of Third Molar Surgery JCShaklin Mustak HussainNo ratings yet
- Alternative techniques for failure of conventional inferior alveolarDocument20 pagesAlternative techniques for failure of conventional inferior alveolarShaklin Mustak HussainNo ratings yet
- Classification of impacted teethDocument45 pagesClassification of impacted teethShaklin Mustak HussainNo ratings yet
- INTRODUCTION TO FRACTUREDocument32 pagesINTRODUCTION TO FRACTUREShaklin Mustak HussainNo ratings yet
- sutureDocument23 pagessutureShaklin Mustak HussainNo ratings yet
- The Pituitary GlandDocument24 pagesThe Pituitary GlandShaklin Mustak HussainNo ratings yet
- Tmj AnatomyDocument29 pagesTmj AnatomyShaklin Mustak HussainNo ratings yet
- INTRODUCTION OF IMPACTIONDocument31 pagesINTRODUCTION OF IMPACTIONShaklin Mustak HussainNo ratings yet
- A Review of The Applications of The Hydrofiber Dressing With Silver Aquacel Ag in Wound CareDocument8 pagesA Review of The Applications of The Hydrofiber Dressing With Silver Aquacel Ag in Wound CareIsabella GeovanaNo ratings yet
- DR Harikrishna - Modern Wound CareDocument147 pagesDR Harikrishna - Modern Wound CareAdrian Lim100% (18)
- AY22 - NUR101 - HB1 - Tutorial 3Document41 pagesAY22 - NUR101 - HB1 - Tutorial 3Siena YipNo ratings yet
- Silver in Wound TherapyDocument28 pagesSilver in Wound TherapyGreg Wilby100% (1)
- Muhs Bhalani 2yrDocument53 pagesMuhs Bhalani 2yrsanjanajaiswal14503100% (2)
- WOUND HEALING ACTIVITY OF METHONOLIC EXTRACT OF Martynia Annua L. (MARTYNIACEAE)Document5 pagesWOUND HEALING ACTIVITY OF METHONOLIC EXTRACT OF Martynia Annua L. (MARTYNIACEAE)xiuhtlaltzinNo ratings yet
- BNM Light Therapy Device Presentation LampDocument10 pagesBNM Light Therapy Device Presentation LampBrane StankovicNo ratings yet
- Pharmaceutics 14 00991 v2Document38 pagesPharmaceutics 14 00991 v2ela.sofiaNo ratings yet
- Review On Trichosanthes DioicaDocument8 pagesReview On Trichosanthes DioicaDevansh MehtaNo ratings yet
- Tissue EngineeringDocument17 pagesTissue EngineeringAntonio LoureiroNo ratings yet
- Wound Healing and Its Impairment in The Diabetic Foot: ReviewDocument9 pagesWound Healing and Its Impairment in The Diabetic Foot: ReviewJoey TsaiNo ratings yet
- Fish DiseasesDocument811 pagesFish DiseasesAurea VerasNo ratings yet
- Basic Principles of Wound Management - UpToDateDocument32 pagesBasic Principles of Wound Management - UpToDateAndrea RezabalaNo ratings yet
- Osmosis Integumentary SystemDocument6 pagesOsmosis Integumentary SystemVianna CuevasNo ratings yet
- WoundDocument45 pagesWoundHailemariam MebratuNo ratings yet
- TargetingDocument13 pagesTargetingpaulo saNo ratings yet
- 10-Tissue RepairDocument75 pages10-Tissue Repairraanja2No ratings yet
- Scratch Wound Healing Assay: Simona Martinotti and Elia RanzatoDocument5 pagesScratch Wound Healing Assay: Simona Martinotti and Elia RanzatoNeha MasarkarNo ratings yet
- Evidence-Based Medicine: Wound ManagementDocument16 pagesEvidence-Based Medicine: Wound ManagementRafael FerreiraNo ratings yet
- Astringent Herbs Stambhana KarmaDocument4 pagesAstringent Herbs Stambhana Karmaqueencel100% (1)
- Bu VonnyDocument41 pagesBu VonnyZola IsmuNo ratings yet
- Fundamentals of Nursing - by Darius CandelarioDocument332 pagesFundamentals of Nursing - by Darius CandelarioEmman RamosNo ratings yet
- How To... Ten Top Questions and Answers On The Use of Dressings For Pressure Ulcer PreventionDocument32 pagesHow To... Ten Top Questions and Answers On The Use of Dressings For Pressure Ulcer Preventionelviana kabanNo ratings yet
- Lactobacillus reuteri extracts promoted wound healing via PI3K/AKT/ β-catenin/ TGF β1 pathwayDocument11 pagesLactobacillus reuteri extracts promoted wound healing via PI3K/AKT/ β-catenin/ TGF β1 pathwayYuniarNo ratings yet
- Basic Surgical SkillDocument19 pagesBasic Surgical Skillshadhana sivakumarNo ratings yet
- ML of Injuries-2Document29 pagesML of Injuries-2GreeshmaNo ratings yet
- Lecture NotesDocument5 pagesLecture NotesErnest Patrick MatiasNo ratings yet
- Ent Mcqs For Part I Exam: Prepared By: Dr. Fouad ShamsanDocument86 pagesEnt Mcqs For Part I Exam: Prepared By: Dr. Fouad ShamsanAli Quwarah100% (1)