Per Nordin  ,
Erika Olofsson,
Karin Hjelm
,
Erika Olofsson,
Karin Hjelm
Within-site adaptation: growth and mortality of Norway spruce, Scots pine and silver birch seedlings in different planting positions across a soil moisture gradient
Nordin P., Olofsson E., Hjelm K. (2023). Within-site adaptation: growth and mortality of Norway spruce, Scots pine and silver birch seedlings in different planting positions across a soil moisture gradient. Silva Fennica vol. 57 no. 3 article id 23004. https://doi.org/10.14214/sf.23004
Highlights
- A soil moisture map could be used to support the choice of planting position for different soil moisture conditions
- Mounds reduced mortality rates for conifers when conditions were wet, but at drier conditions differences between planting positions were small
- Contradictory, silver birch had higher survival in lower planting positions compared with mounds
- Height and diameter were higher in mounds for conifers, but only small differences occurred between planting positions for silver birch.
Abstract
Adapting to site conditions is a central part of forest regeneration and can be done through selection of different planting positions. Requirements are tree species specific, and the use of soil moisture maps could be a way to support decision making in forest regeneration planning. At two experimental sites with varying soil moisture conditions in southern Sweden Norway spruce (Picea abies (L.) Karst.), Scots pine (Pinus sylvestris L.), and silver birch (Betula pendula Roth) seedlings were planted in four different planting positions following mounding site preparation; Depression, Hinge, Mound and Unscarified. Soil moisture estimates were obtained from a high-resolution depth-to-water raster for each planting spot. The effect of soil moisture, planting position and their interactions on mortality, height and diameter was evaluated for each tree species. In wet conditions mounds proved to be the best option to minimize seedling mortality for conifers, but with decreasing soil moisture, differences between the planting positions decreased. Birch on the other hand had the greatest survival in the hinge. The coniferous species displayed increased height and diameter when planted in mounds independent of the soil moisture conditions, whereas silver birch was less dependent on a specific planting position. Results from this study shows that a soil moisture map can explain mortality, height and diameter and thus can be a useful tool when choosing planting position in different soil moisture conditions.
Keywords
regeneration;
microsite;
decision making;
mounding;
planting;
depth-to-water
-
Nordin,
pcSKOG, Grisslevägen 15, 227 32 Lund, Sweden
 https://orcid.org/0000-0002-2156-6615
E-mail
per.nordin@skogforsk.se
https://orcid.org/0000-0002-2156-6615
E-mail
per.nordin@skogforsk.se
-
Olofsson,
Linnaeus University, Department of Forestry and Wood Technology, 351 95 Växjö, Sweden
https://orcid.org/0000-0001-5844-6775
E-mail
erika.olofsson@lnu.se
-
Hjelm,
Swedish University of Agricultural Sciences, Southern Swedish Forest Research Centre, Box 190, 234 22 Lomma, Sweden
https://orcid.org/0000-0002-6144-8250
E-mail
karin.hjelm@slu.se
Received 23 January 2023 Accepted 23 August 2023 Published 14 September 2023
Views 16909
Available at https://doi.org/10.14214/sf.23004 | Download PDF

1 Introduction
Adaptation to a site is often discussed as crucial to succeed with regeneration and to fulfill the full growth potential (Albrektsson et al. 2012). Choosing tree species and selecting suitable management options is a way to adapt to site conditions. Tree species respond differently to various stress factors, making them favorable in certain environmental conditions, and various management options are more or less appliable to ameliorate site conditions (Nilsson et al. 2010; Wiensczyk et al. 2011; Löf et al. 2012; Aleksandrowicz-Trzcińska et al. 2017; European and Joint Research 2022). Adaptations to site conditions are often done on a site level, even though most sites show variation on a microsite level regarding soil type, soil moisture and other important factors. This has raised the awareness of microsite adaptation to reach higher precision in silvicultural practice (Holopainen et al. 2014; Fardusi et al. 2017; Saksa et al. 2021; Persson et al. 2022).
The choice of planting spot can be seen as site adaptation on a microsite level where different local site properties can be suitable for the planted seedlings to a varying degree (Luoranen et al. 2018; Häggström et al. 2021; Nordin et al. 2022). The environment on a clear-cut leads to significant seedling stress (Margolis and Brand 1990) emphasizing the need to choose a suitable planting spot aiming at fast seedling establishment (Örlander et al. 1990; Thiffault and Jobidon 2006; Marquis et al. 2021). Following mechanical site preparation, several planting spots with diverse microsite conditions are created (Löf et al. 2012; Wallertz et al. 2018; Sikström et al. 2020a). On elevated planting spots, soil temperature and hydrological conditions are different from lower spots (Sutton 1993). In berms and capped mounds, the porous structure with humus underneath a mineral soil cap could increase the risk of drying when precipitation is low, thus limiting water availability (Luoranen et al. 2018; Häggström et al. 2021). In contrast, lower planting positions in humid environments could increase the risk of oxygen deficiency, due to the accumulation of water, but be more favorable in dry environments (Örlander et al. 1990; Hansson et al. 2018). Nutrient availability is also important to consider since mounds and planting spots on top of humus has shown to promote mineralization (Schmidt et al. 1996; Smolander and Heiskanen 2007), and planting spots where humus has been removed can lead to nutrient deficiency (Nohrstedt 2000; Simard et al. 2003). This can have a significant impact on Norway spruce (Picea abies (L.) Karst.) seedlings that have shown to be more sensitive to removal of nutrients compared to Scots pine (Pinus sylvestris L.) (Nilsson et al. 2019). Also, planting spots with certain characteristics may reduce abiotic and biotic risks such as frost or pine weevil (Hylobius abietis L.) (Langvall et al. 2001; Nordlander et al. 2011; Heiskanen et al. 2013; Luoranen et al. 2017).
Recommendations regarding the selection of planting spots seldom differ between tree species in Sweden (Albrektsson et al. 2012). However, Scots pine and Norway spruce have been shown to respond differently to site preparation (Nilsson et al. 2019). Also, there is limited knowledge about planting of birch (Betula L. spp.) on forest land since general practice and research has focused on natural regeneration (Perälä and Alm 1990; Karlsson and Nilsson 2005; Hynynen et al. 2010; Holmström et al. 2017; Saursaunet et al. 2018) or planting of silver birch (Betula pendula Roth) on former agriculture land (Karlsson 2002; Daugaviete et al. 2003; Hytönen and Jylhä 2005; Rytter and Lutter 2020). Only a few studies have examined the effects of different planting spots on forest land (Luoranen et al. 2003; Pikkarainen et al. 2021). Present knowledge has led to recommendations for planting of silver birch on forest land that are often similar to those for conifers (Rytter et al. 2014; Fahlvik et al. 2021), but with an emphasis on the need of vegetation control (Karlsson 2002; Hytönen and Jylhä 2013).
Within-site variation of soil moisture can lead to differing results in growth and mortality of seedlings and motivate a variation in management (Skovsgaard and Vanclay 2013; Holmström et al. 2019). Through advancements in remote sensing, digital soil moisture maps of high resolution are available, making it easier to detect within-site variation and assist in decision making (Murphy et al. 2008; Ågren et al. 2014; Nijland et al. 2015). Soil moisture maps have provided support in practical forest management planning to avoid rutting damage made by forest machinery (Mohtashami et al. 2017), to protect forest streams and plan buffer zones (Kuglerová et al. 2014; Ågren et al. 2015), or as source of recommendations regarding site preparation (Ring et al. 2020). There is also potential to use these maps for regeneration planning to reduce seedling mortality (Holmström et al. 2019). Further development of these maps, combined with site knowledge, could enable splitting of clear-cuts into smaller homogenous areas for potential uses including tree species selection (Saksa et al. 2021). The evolution of digital tools could therefore be a stepping stone towards precision forestry with more detailed information available for decision making.
The purpose of this study was to study the effect of soil moisture, planting position and their interaction on mortality, height and diameter of Norway spruce, Scots pine and silver birch seedlings. Two sites with great within-site variation in soil moisture were used, which enabled comparisons between different soil moisture conditions under the same climatic conditions. For evaluation of soil moisture, a high-resolution soil moisture map was used to enable evaluation on a microsite level. We therefore also evaluate if a soil moisture map could be used for decision making regarding which planting position to choose in different conditions. We hypothesize that:
1) On wet sites, elevated planting positions (mounds) increase seedling survival
2) On drier sites, planting positions will be of less importance for survival
3) Seedling height and diameter will be highest in elevated mounds
2 Material and method
2.1 Experimental design
Two sites in southern Sweden, Isberga (Isb) in the province of Småland and Holkaberga (Holk) in the province of Skåne, were selected for establishment of the experiment (Fig. 1). They were selected due to their within-site variation in soil moisture, which was essential for the purpose of exploring the use of soil moisture maps for regeneration planning. The sites were clear-felled in 2014 (Isb) and 2018 (Holk) with subsequent site preparation using an excavator in 2019 (Isb) and 2020 (Holk) resulting in mounds. Additionally, a ditch cleaning was carried out in the spring prior to establishment in Holk. The sites differed in site characteristics such as clear-cut ages, site index and climatic conditions (Table 1). Soil texture at both sites were predominantly sandy till soil but differed with soil moisture conditions. In Isb the wetter parts were dominated by peat, with a thick humus layer. Historical maps (Lantmäteriet 2022) indicated that the wetter parts of Holk, with more organic material mixed in the soil, were possibly pasture or farmland in the past.
Fig. 1. Map displaying the location of the two experimental sites Isberga (filled triangle) in the province of Småland and Holkaberga (filled dot) in the province of Skåne where this study was conducted.
| Table 1. Site characteristics of the two experimental sites in southern Sweden used to study the effect of planting position and soil moisture conditions on the three species Norway spruce, Scots pine and silver birch. | ||
| Site | Isberga | Holkaberga |
| Year of clear felling | 2014 | 2018 |
| Year of establishment a | 2019 | 2020 |
| Site index b | G24 | G34 |
| Coordinates (WGS84) | 57°13´N, 13°25´E | 56°01´N, 13°50´E |
| Elevation (m a.s.l.) | 163 | 115 |
| Precipitation (mm) c | ||
| 1991–2020 | 983 (587) | 843 (486) |
| Study period | 1020 (567) | 844 (463) |
| Temperature (°C) d | ||
| 1991–2020 | 6.6 (11.4) | 8.1 (12.7) |
| Study period | 7.7 (12.0) | 9 (13.3) |
| Norway spruce e | SP75 / Lilla Istad FP – 502 | SA115 / Ekebo FT – 907 |
| Scots pine e | SA90 / Lilla Istad FP – 604A | SP75 / Gotthardsberg FP – 606B |
| Silver birch e | BP36 / Ekebo 5 FP – 809 | Plantek36 / Ekebo 5 FP – 809 |
| a Planting was done in the same year as the experimental establishment. b Site index derived from site properties as the height of the dominant trees after 100 years for Norway Spruce (G). c Average annual precipitation, derived from the nearest weather station (SMHI 2022) for the latest climate period (1991–2020) and the study period (2019–2021 Isberga and 2020–2022 Holkaberga). The value within the brackets represents the average precipitation during the growing season (April–October). Weather stations; Isberga – Skeppshult D (73070) (57°12´N, 13°40´E) and Holkaberga – Knopparp D (53570) (55°93´N, 13°85´E). d Average mean air temperature derived from the nearest weather station (SMHI 2022) for the latest climate period (1991–2020) and the study period (2019–2021 Isberga and 2020–2022 Holkaberga). Values within the brackets represent the average mean air temperature during the growing season (April–October). Weather stations: Isberga – Hagshult (74180) (57°29´N, 14°14´E) and Holkaberga – Hörby A (53530) (55°86´N, 13°67´E). e Information about the seedling type, i.e. container type (e.g. SP75) and provenance of the seed source (e.g. Lilla Istad FP – 502). Container type and provenance of the seed source are separated with a dash. | ||
To ensure that the sites included a wide range of soil moisture conditions, a depth-to-water (DTW)-raster (Ågren et al. 2014; Murphy et al. 2008) was used when establishing the experiment. For each site, 25 blocks were established evenly distributed across four different soil moisture conditions, wet, moist, mesic and dry (Fig. 2). In each block, 12 mounds were created with an excavator. Because of the within-site variation, the mounds differed in size, resulting in differences in the depth and height of the planting positions. Seedlings of the same species were planted in four different planting positions in the same mound: (1) depression, (2) hinge, (3) mound, and (4) unscarified control (Fig. 3). The height below and above ground of each planting position was measured in the autumn after planting. On average, seedlings were planted 22 cm below ground surface in depression, 11 cm below ground in hinge and 12 cm above ground in mound. The surface of the planting spots (i.e. mineral soil) was around 30 cm in diameter.
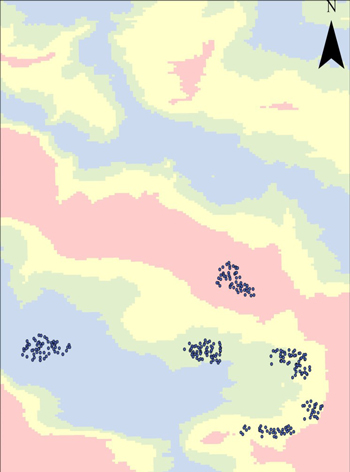
Fig. 2. Distribution of the planting spots in the different soil moisture conditions according to the soil moisture map (Depth-to-water-raster) at the experimental site Isberga in the province of Småland. Blue color = wet soil (superficial DTW, 0–0.5 m), green = moist (intermediate, 0.51–1 m), yellow = mesic (deep DTW, 1.01–2 m) and pink = dry (very deep DTW, 2.01 m-infinity). The DTW-raster was downloaded from the Swedish Forest Agency (SFA).
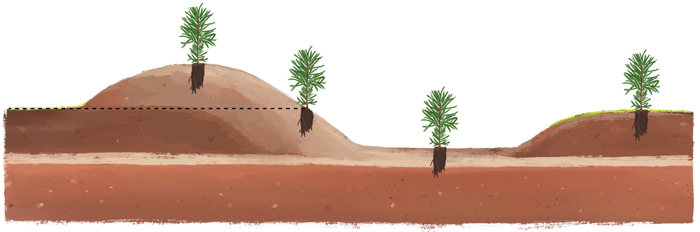
Fig. 3. Illustration of the different planting positions within a planting spot (mound) used in this study to check the effects on seedlings of Norway spruce, Scots pine and silver birch in different soil moisture conditions. From the left; Mound, Hinge, Depression, and Unscarified.
Each block was planted with 48 seedlings, 16 seedlings per species adding up to a total of 1200 seedlings per site. Containerized improved seedling material of Norway spruce, Scots pine and silver birch provided by Svenska Skogsplantor was used (Table 1). The seedling material, i.e. provenances, was selected to fit the climatic conditions of the two sites. All seedlings were planted at a conventional planting depth, i.e. where the peat plug was buried a couple of centimeters below the soil surface. Silver birch seedlings were generally larger than both the Norway spruce and Scots pine seedlings. The silver birch seedlings were thus planted using a planting auger, while the Norway spruce and Scots pine seedlings were planted with a planting tube. If standing water was present in the depression, the seedling was planted as low as possible without being submerged. Norway spruce and Scots pine seedlings were treated with a Conniflex coating (Svenska Skogsplantor), a composition of sand and glue applied onto the stem of the seedlings in the nursery before delivery, to protect against pine weevil damage. To handle potential browsing, all seedlings were treated with Trico (Organox AB), an emulsion of fatty acids from sheep that is sprayed on the leading shoot, in the autumn after the first and second growing seasons.
2.2 Field measurements
Seedling height (in mm) above soil surface and diameter at ground level (in mm) was measured after each one of the first three growing seasons. Also, the damage level of the seedlings was assessed at the same time using a 7-level scale: 0= No damage, 1 = Negligible damage, 2 = Slightly damaged (reduced growth but not smaller than previous year), 3 = Severe damage (smaller than previous year), 4 = Lethal damage (expected to die the following year), 5 = Dead, and 6 = Missing. The major cause of damage was also registered, such as browsing, damage by insects (mostly pine weevil), water logging or fungi-related damage. When the cause of damage could not be determined, it was registered as unknown. Only the major cause of damage was registered per seedling at each inventory occasion.
Soil moisture content was measured using a TDR (Time-domain reflectometer, FieldScout TDR 350 Soil Moisture Meter) at a depth of 0.2 m. Measurements were taken in all planting positions in one random mound per block and site once a month from May to October during the two first growing seasons in Isb and the first and third growing season in Holk. The average soil moisture content was then calculated for each site to represent the actual soil moisture condition during the growing season.
2.3 Digital data
To spatially identify each individual planting spot, they were positioned using the Avenza maps application for smartphones (Avenza MapsTM). The built-in smart phone GNSS (Global Navigation Satellite Systems) had an accuracy of ±2 m. Soil moisture values for each planting spot were extracted from a national DTW-raster (2 × 2 m resolution) available at the Swedish Forest Agency (SFA 2022) using ArcGIS Pro (Esri, West Redlands, CA). A DTW-raster displays the estimated depth to groundwater as calculated based on the topographical variation and proximity of water sources with a predetermined stream initiation threshold (Murphy et al. 2008; Ågren et al. 2014). Values of the DTW-raster can range between 0m to 1000 m, where values closer to 0indicate wetter conditions. The distribution of the DTW-values in relation to soil moisture was not linear, meaning that differences in soil moisture for values between 0–1 m are greater than values from 1 m and upwards. Therefore, a classification of the values was needed before analysis, a procedure also made in previous studies using DTW (Ågren et al. 2015; Mohtashami et al. 2022). The values were classified into four classes: Superficial DTW, 0–0.5 m, Intermediate DTW, 0.51–1 m, Deep DTW, 1.01–2 m, Very deep DTW, 2.01 m–infinity. In practice, these are mainly referred to as wet, moist, mesic and dry, respectively.
2.4 Data analysis
Mortality was determined as the amount of seedlings within vitality class 5 and 6 after three growing seasons. To analyze the effect of DTW and planting position on mortality, a mixed generalized linear model with a binomial distribution was constructed using Proc Glimmix in SAS 9.4 (SAS Institute, Cary, NC, USA). The following model was fitted:
![]()
where µ is the overall mean, αi the fixed effect of DTW class (i = 1–4), bij is the random effect of DTW class within site, γk is the fixed effect of planting position (k = 1–4), (αγ)ik is the interaction between the fixed effects DTW class and planting position, δl is the fixed effect of tree species (l = 1–3), (αδ)il is the interaction of the fixed effects DTW class and tree species, (γδ)kl is the interaction between the fixed effects planting position and tree species, (αγδ)ikl is the three-way interaction between the fixed effects DTW class, planting position and, tree species and εijkl is the experimental error. If significance was detected, a Tukey adjusted pairwise comparison was conducted to differentiate between lsmeans of DTW-class, planting position and species.
A linear mixed effect model was used to analyze height and diameter after three growing seasons. Since growth patterns naturally differ between species and therefore might be difficult to compare in an analysis, height and diameter were analyzed separately for each species. Both height and diameter followed a normal distribution, and the following model was fitted with Proc Mixed in SAS 9.4 (SAS Institute, Cary, NC, USA):
![]()
where µ is the overall mean, αi the fixed effect of DTW class (i = 1–4), bij is the random effect of DTW class within site, γk is the fixed effect of planting position (k = 1–4), (αγ)ik is the interaction between the fixed effects DTW class and planting position and εijk is the experimental error. The denominator degrees of freedom were calculated using the Satterthwaite method. A Tukey adjusted pairwise comparison between lsmeans was conducted when variables were significant, and the significance level was set to 0.05.
3 Results
3.1 Soil moisture
For both sites, measured soil moisture content and DTW correlated in the unscarified control planting position (Fig. 4). When DTW increased, the soil moisture content decreased, indicating that DTW sufficiently captured the differences in soil moisture at both sites. As expected, depression and hinge had the highest soil moisture content and the mound the lowest with superficial and intermediate DTW (Fig. 5). However, when DTW increased (deep and very deep) and indicated drier conditions, the soil moisture content in the hinge and depression decreased and became similar to mound and unscarified. Notably, soil moisture content was generally lower in Holk compared to Isb.
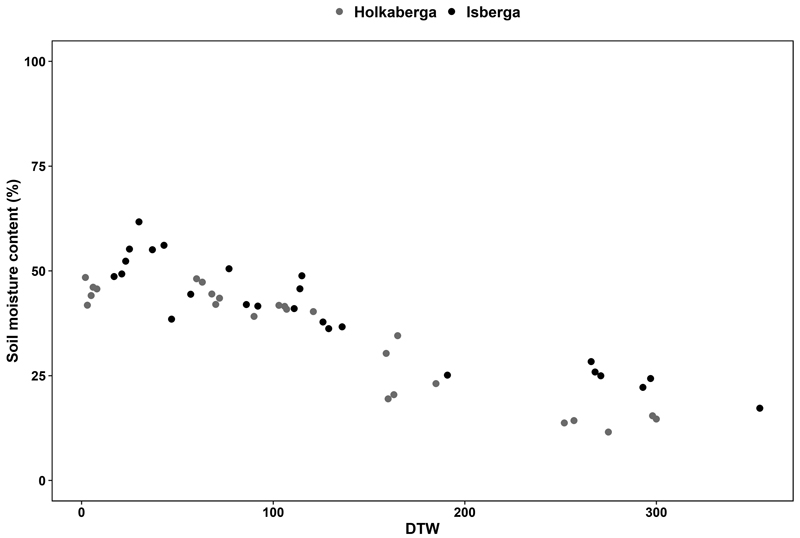
Fig. 4. The relation between depth-to-water (DTW), in cm, and soil moisture content (%) measured with Time domain reflectometer (TDR) in the unscarified control once a month between May and October for two years at the experimental site in Holkaberga (gray dots) and Isberga (black dots) in southern Sweden. Only measurements in the unscarified control are shown in this graph to represent the conditions when soil conditions have been unaltered.
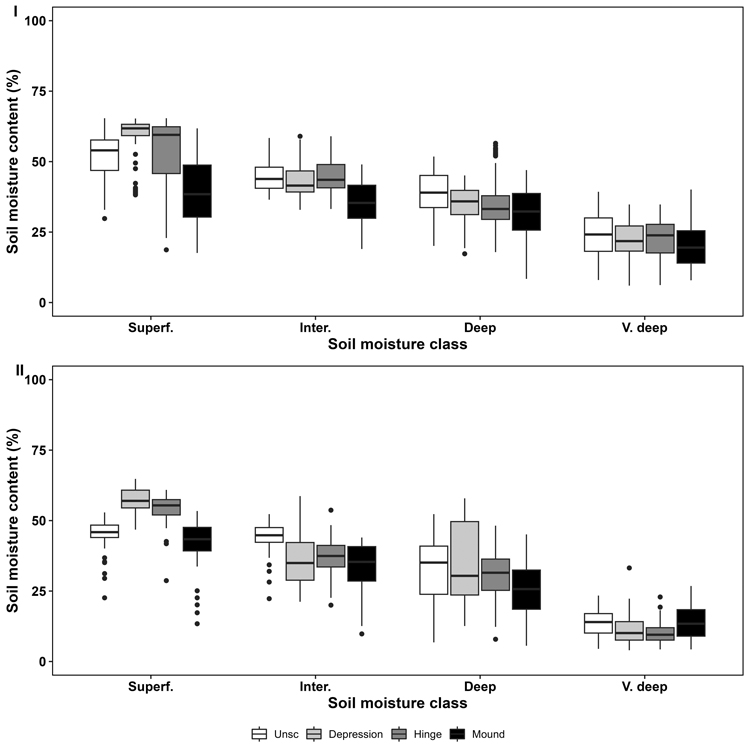
Fig. 5. Soil moisture content measured using a Time domain reflectometer (TDR), during two growing seasons in May to October in relation to soil moisture derived from the depth-to-water (DTW)-map in the four planting positions for the two experimental sites Isberga (I) and Holkaberga (II). Superf. = Superficial DTW (0–0.5 m depth); Inter. = Intermediate DTW (0.51–1 m depth); Deep = Deep DTW (1.01–2 m depth); V. deep = Very deep DTW (2.01 m – infinity depth). UnSc = Unscarified (white); Depression (light gray); Hinge (gray); Mound (black).
3.2 Mortality
There was a significant interaction between DTW class, planting position and species for mortality (Table 2). The greatest effect was found for Norway spruce, where planting in depression in wet soil conditions (superficial DTW) generated the highest mortality with 96% dead seedlings, in contrast to the mound which achieved the lowest value with 4% (Fig. 6). As soil moisture conditions became drier (increasing DTW depth), the planting positions became less important, but mortality in depression tended to be higher. In very deep DTW, mortality was similar in all planting spots, around 15%. Instead, here the unscarified position generated the highest mortality of 47%.
| Table 2. Results of the analysis of variance (p-values) for the fixed effects and their interaction after the first three growing seasons on mortality for the species Norway spruce, Scots pine and silver birch. Statistically significant effect (<0.05) is highlighted in bold. DTW = depth-to-water, Plantpos = planting position. | |
| p-value | |
| Effect | Mortality |
| DTW class | 0.3590 |
| Plantpos | <0.0001 |
| DTW class × Plantpos | <0.0001 |
| Species | 0.1152 |
| DTW class × Species | 0.4700 |
| Plantpos × Species | <0.0001 |
| DTW class × Plantpos × Species | 0.0144 |
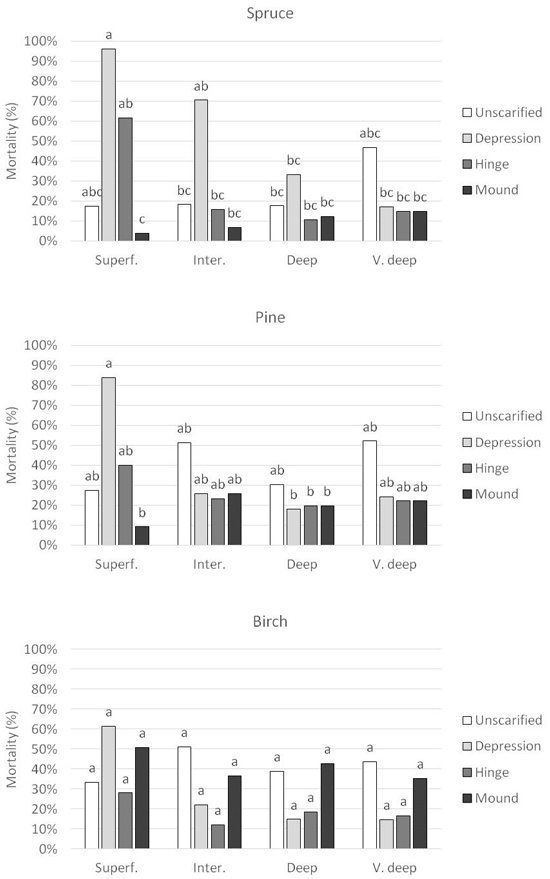
Fig. 6. Seedling mortality for the respective species, Norway spruce, Scots pine and silver birch planted in the four planting positions in each Depth-to-water (DTW) class. DTW classes are Superficial DTW (0–0.5 m depth), Intermediate DTW (0.51–1 m depth), Deep DTW (1.01–2 m depth), and Very deep DTW (2.01 m – infinity depth). Superf. = Superficial DTW; Inter = Intermediate DTW; Deep = Deep DTW; V. deep = Very deep DTW. Columns with different letters are significantly different. Since significant interactions occurred, comparisons are made between both DTW class and planting positions.
For Scots pine, the effect of DTW class and planting spot was not as pronounced as for Norway spruce (Fig. 6). Still, in superficial DTW, the differences were large with 84% mortality in depression compared with 9% in the mound. In the other DTW-classes, there were no differences between the planting positions, and the mortality varied around 20%. Here, the unscarified position in general generated the highest mortality between 30–50%.
No significant differences for DTW class and planting position were found for silver birch (Fig. 6). Although not significant, the highest mortality was found in the depression in superficial DTW with 61%. In contrast to Norway spruce and Scots pine, more seedlings tended to die in the mound and unscarified position in intermediate, deep and very deep DTW, 35–50%, compared to 12–20% in depression and hinge.
As a summary, the overall best planting position for Norway spruce was the mound (8% mortality) and the worst was depression (54% mortality). For Scots pine, mound was most preferred (19% mortality) and depression least preferred (39% mortality). Birch performed best in hinge (20% mortality) and worst in mound (42% mortality).
The major cause of mortality differed between planting spots and species (Table 3). The main factor resulting in mortality for seedlings planted in depression and hinge was water logging which could have caused oxygen deficiency. For Norway spruce, insect damage, primarily pine weevils, was the dominant factor causing mortality. For Scots pine, browsing and insect damage dominated, while for silver birch fungi (stem spot lesion) and browsing were the major reasons.
| Table 3. Major cause of damage for each species and planting spot treatment in percent of total damage for all seedlings used in the study. The percentage represents the combined results from both experimental sites Isberga and Holkaberga. The major causes of damage are represented by drought, water logging, fungal damage, browsing damage, insect damage, and unknown cause of damage. | |||||||
| Drought | Water a | Fungi | Browsing | Insects | Unknown | ||
| Spruce | Unscarified | 2 | 8 | 39 | 51 | ||
| Depression | 4 | 70 | 1 | 12 | 13 | ||
| Hinge | 2 | 62 | 13 | 23 | |||
| Mound | 21 | 63 | 16 | ||||
| Pine | Unscarified | 7 | 1 | 15 | 28 | 49 | |
| Depression | 59 | 4 | 11 | 26 | |||
| Hinge | 2 | 26 | 8 | 25 | 39 | ||
| Mound | 27 | 16 | 57 | ||||
| Birch | Unscarified | 43 | 40 | 4 | 13 | ||
| Depression | 36 | 29 | 12 | 23 | |||
| Hinge | 10 | 44 | 26 | 20 | |||
| Mound | 36 | 38 | 1 | 25 | |||
| a Water = Water logging. | |||||||
3.3 Height
Significant interactions between DTW class and planting position occurred for all three tree species regarding height after three growing seasons (Table 4). For Norway spruce, this interaction showed that the greatest difference in height between planting positions was found in superficial DTW (Fig. 7). Seedlings grown in mounds achieved the highest height, 765 mm, and this was significantly higher than seedlings grown in depression, 445 mm. In the other DTW classes, no significant differences were found between planting spots. A similar pattern was found for Scots pine, where the height in mounds in superficial DTW were significantly higher than in the depression, 765 mm versus 448 mm (Fig. 7). For Scots pine, seedlings planted in the unscarified treatment were also significantly higher, 756 mm, compared with the depression. No other differences were found, but the unscarified treatment tended to have seedlings with lower height in intermediate, deep and very deep DTW. For silver birch, the only differences found between planting spots were in deep DTW (Fig. 7). Here, height for seedlings in mound were significantly lower than for seedlings planted in hinge or depression, 799 mm compared with 1117 mm and 1138 mm, respectively. The same pattern was found for intermediate and very deep DTW, but not significant.
| Table 4. Results of the analysis of variance (p-values) for the fixed effects and their interaction after three growing seasons on height and diameter for Norway spruce, Scots pine, and silver birch. Each species was analyzed separately. Statistically significant effect (<0.05) is highlighted in bold. DTW = depth-to-water and Plantpos = planting position. | |||
| p-value | |||
| Species | Effect | Height | Diameter |
| Norway spruce | DTW class | 0.4819 | 0.3997 |
| Plantpos | 0.0003 | <0.001 | |
| DTW class × Plantpos | 0.0026 | <0.001 | |
| Scots pine | DTW class | 0.8093 | 0.7383 |
| Plantpos | 0.0140 | <0.001 | |
| DTW class × Plantpos | <0.001 | 0.0015 | |
| Silver birch | DTW class | 0.8267 | 0.7915 |
| Plantpos | <0.0001 | 0.0513 | |
| DTW class × Plantpos | 0.0106 | 0.0003 | |
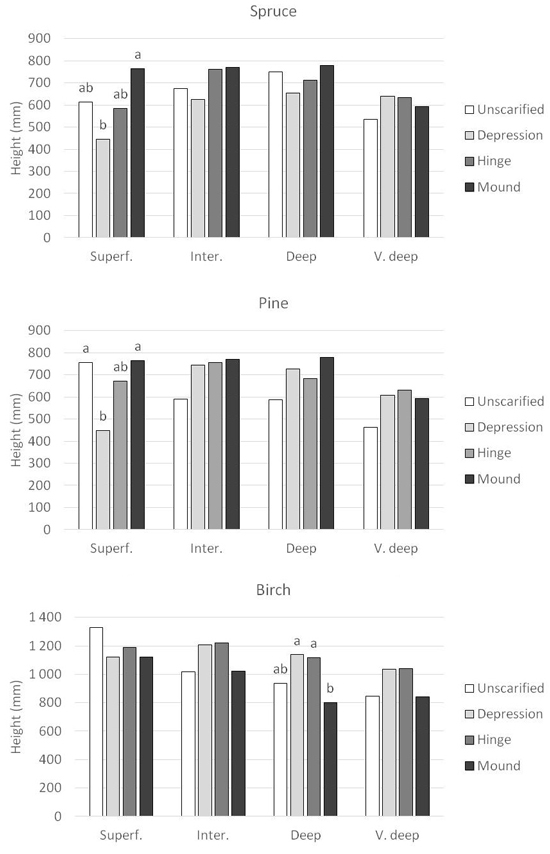
Fig. 7. Seedling height (mm) for the respective species, Norway spruce, Scots pine and silver birch planted in the four planning positions in each Depth-to-water (DTW) class. DTW classes are Superficial DTW (0–0.5 m depth), Intermediate DTW (0.51–1 m depth), Deep DTW (1.01–2 m depth), and Very deep DTW (2.01 m – infinity depth). Superf. = Superficial DTW; Inter = Intermediate DTW; Deep = Deep DTW; V. deep = Very deep DTW. Columns with different letters are significantly different within each DTW class. Columns without letters are not significantly different. Notice the different scale on the Y-axis for silver birch compared to Norway spruce and Scots pine.
3.4 Diameter
As for height, significant interactions between DTW class and planting position occurred for all three tree species after three growing seasons (Table 4). For Norway spruce, seedling diameter was significantly highest in mounds in superficial, intermediate and deep DTW (Fig. 8). The difference was greatest in superficial DTW with a diameter of 17 mm in the mounds compared with around 9–11 mm in unscarified, depression and hinge. The differences declined with increasing DTW class and were not significant in very deep DTW. For Scots pine, seedlings planted in mounds also had the highest diameter in general (Fig. 8). As for Norway spruce, the difference between planting positions was most pronounced in superficial DTW where the diameter in mounds was 21 mm while it was only 8 mm in the depression. In intermediate, deep and very deep DTW, diameter in depression was similar to unscarified and hinge. Less variation in diameter was found for silver birch (Fig 8). No significant differences were found between DTW class and planting position except for in superficial DTW where seedling in unscarifed positions had a significantly higher diameter than in depression, 18.5 mm versus 14 mm.
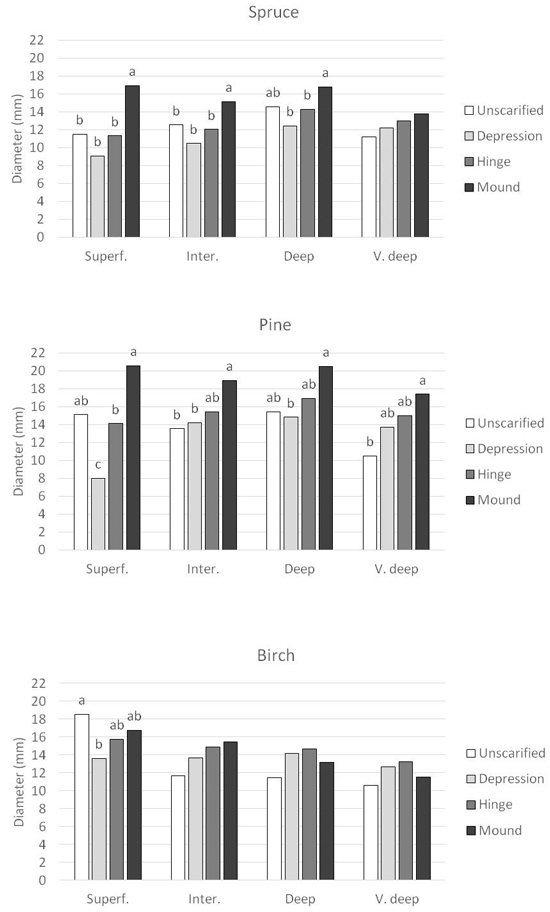
Fig. 8. Seedling diameter (mm) for the respective species, Norway spruce, Scots pine and silver birch planted in the four planning positions in each Depth-to-water class. DTW classes are Superficial DTW (0–0.5 m depth), Intermediate DTW (0.51–1 m depth), Deep DTW (1.01–2 m depth), and Very deep DTW (2.01 m – infinity depth). Superf. = Superficial DTW; Inter = Intermediate DTW; Deep = Deep DTW; V. deep = Very deep DTW. Columns with different letters are significantly different within each DTW class. Columns without letters are not significantly different.
4 Discussion
4.1 Mortality
This study showed that soil moisture derived from a DTW-map and planting position after mounding significantly explained the mortality rate of Norway spruce, Scots pine and silver birch, to varying degrees, on two forest sites in southern Sweden. As could be expected, the variation in mortality within areas of different soil moisture conditions varied largely depending on the planting position. Notably, it was the wettest soil moisture class that displayed the largest variation between planting positions regarding mortality for the conifers. When conditions were wetter, a lot of the mortality was due to seedlings planted in the depression. High mortality rates in the depression were not a surprise, as waterlogging is a known issue when regenerating in areas with high soil moisture content (Pearson et al. 2011). Seedlings planted below the soil surface in environments with high soil moisture suffer from waterlogging and will either develop poorly or die (Örlander et al. 1990; Grossnickle 2016; Henneb et al. 2019). Hence, in such areas the recommendation is to search for elevated planting spots (Örlander et al. 1990; Sutton 1993), which was also true in this study. Our results support the hypothesis that mounds are suitable options to decrease the mortality rate, particularly for the conifers. When soil moisture conditions become drier, depression and hinge positions prove to be as good options as mounds to reduce the mortality rate. In areas or occasions with low precipitation, low planting positions can even be preferred to avoid mortality due to drought when mounds run the risk of drying out (Örlander et al. 1990; Luoranen et al. 2018; Häggström et al. 2021; Nordin et al. 2022).
Contradictory, silver birch seedlings planted in the elevated mounds had a rather high mortality rate, although not statistically different, it was always higher than in the hinge. Mounding is known to be a viable planting position option in wet areas for both Norway spruce and Scots pine to reduce the mortality rate (Nilsson and Örlander 1995; Johansson et al. 2007; Pearson et al. 2011; Heiskanen et al. 2013). For silver birch, mounding is also a viable option but has been studied to a lesser degree in comparative studies (Pikkarainen et al. 2021). However, other broadleaved tree species, like poplars (Populus spp.) and hybrid aspen (Populus tremula L. × P. tremuloides Michx.), have been shown to respond well to planting in mounds (Mc Carthy et al. 2017; Hjelm and Rytter 2018; Thiffault et al. 2020). Our study showed that the mound might not be the best option regarding survival for birch seedlings. One explanation could be that silver birch is a species that requires a lot of water and that the elevated position in the mound became drier than the hinge and depression. Birch often regenerates naturally on moist to wet parts on a clear-cut area and thrives in moist conditions (Lidman et al. 2023).
This study did not clearly show that unscarified planting positions should be avoided to decrease mortality, although it is a common conclusion from previous studies (Sikström et al. 2020a). Probably, the differences in clear-cut age between the sites most likely played into this, since pine weevil damage was not a factor in the five-year-old clear-cut in Isb. Four years after clear-felling a large amount of the pine weevil population has left the clear-cut and the feeding pressure drops significantly (Örlander et al. 1997; von Sydow 1997). In Holk on the other hand, where the clear-cut was one year old, there was an abundance of pine weevil damage, which is also more likely to be severe in unscarified positions (Petersson et al. 2005; Nordlander et al. 2011). Here, the unscarified positions had higher mortality rates, and it was most obvious when conditions were drier.
It is evident that the different species are exposed to various causes of damage. Water logging was the most important factor causing mortality in Norway spruce and Scots pine in depression and hinge, while it was of less importance for birch. Insect damage, mainly by pine weevil, was the reason behind a large part of the mortality in the unscarified and mound position. In silver birch, stem spot lesion explained around 30–40% of the seedling mortality. Browsing or damage made by herbivores hardly caused any damage in Norway spruce, but was a major factor for both Scots pine and silver birch. These two species are highly attractive by browsers (Bergström and Bergqvist 1997; Månsson et al. 2007), which can be problematic in the regeneration phase if no measures are taken to reduce damage. In conclusion, the species are exposed to different threats, which needs to be considered when choosing site and planting spot since additional measures to prevent damage might be needed, for example pine weevil protection, browsing repellants, fencing, and by also considering stock type and genetic material that are more resistant to a specific damage agent.
4.2 Height and diameter
Depending on planting position, seedlings responded differently to the soil moisture conditions in terms of height and diameter three years after planting. Clear effects of the excessive amount of water could be seen for Norway spruce and Scots pine planted in depression where both height and diameter were reduced. Lack of oxygen due to the wet conditions can hinder the growth since root growth and functioning is inhibited (Örlander et al. 1990; Grossnickle 2000; Grossnickle 2012; Grossnickle and Macdonald 2018). This growth reduction in the wettest conditions for the coniferous species was especially evident when comparing the mound position to lower positions. The coniferous species had overall a superior growth in the mounds, but the same pattern could not be found for silver birch, not even in the wettest parts of the sites. In terms of height, the browsing level on the silver birch could partly explain the lack of differences between treatments.
Norway spruce seedlings in this study consistently showed a greater height in the mound position, indicating that mounding creates favorable conditions no matter the soil moisture conditions. This is even more prevalent when looking at the diameter of Norway spruce and Scots pine seedlings where planting in the mound was the best option, displaying superior diameter in comparison to the other planting positions, no matter the soil moisture conditions. Increased diameter is generally an indication of root development (Grossnickle 2012; Grossnickle and Macdonald 2018), which further confirms that mounding creates favorable conditions for establishment (Hallsby and Örlander 2004). When mounding, the loosening of soil improves nutrient availability and increases soil temperature (Örlander et al. 1990; Sutton 1993; Smolander and Heiskanen 2007). Also, the consistent higher height in the mound position for the two conifers could be an effect of the removal of competing vegetation (Nilsson and Örlander 1999; Thiffaul et al. 2010). Below-ground competition for water and nutrients have previously been shown to be an important factor in establishment (Munson et al. 1993; Nilsson et al. 1996), though in this study competition for water was most likely not the issue except for the driest areas of the sites.
In terms of height and diameter development, a recommended planting position for silver birch is hard to distinguish from our results since seedlings planted in the unscarified position consistently grew as good as the other planting positions. In fact, contrary to the conifers, the mound position seemed to generate the lowest height and diameter compared to the other treatments, although not always statistically significant. One recurring theme in previous studies on silver birch regeneration research is the need to manage the competing vegetation (Karlsson 2002; Hytönen and Jylhä 2005). Most of the studies have focused on planting on former agricultural land where serious damage by voles and a dense vegetation cover is to be expected (Karlsson 2002; Daugaviete et al. 2003; Hytönen and Jylhä 2005). In this study, forest land was planted, and no such damage was recorded at either site but damage by ungulates seriously hindered the height growth, which may be a common issue when planting birch (Hynynen et al. 2010). The potential lack of enough soil water could be another aspect in the mounding positions. In capped mounds, drought could occasionally be a problem (Häggström et al. 2021).
4.3 Soil moisture
Since the soil moisture derived from the DTW-map showed such an important effect in the analysis, it suggests that there is a use for soil moisture maps in regeneration planning to some extent. Being able to differentiate between areas where elevated planting positions should be recommended is one application. This has previously been suggested as a utilization of soil moisture maps in Swedish forestry, or at least using them to identify areas where mounding is a suitable site preparation method (Ring et al. 2020). The complexity of modelling soil moisture became evident from these two experiments. Local conditions at Isb and Holk played a role in how soil moisture varied locally, which is not always captured by national soil moisture maps like the DTW-map used in this study. Previous validations of different types of soil moisture maps firmly confirms that for the most accurate result, tuning of resolution and threshold values should be done for the area of interest (Larson et al. 2022). However, as a planning tool, the national maps are viable in terms of dividing and identifying different areas where special management is needed, which has a great value in forest regeneration planning (Holmström et al. 2019).
4.4 Conclusion
Our results show that mortality, height and diameter of Norway spruce, Scots pine and silver birch could be improved by using a soil moisture map to select a planting position suitable for a given condition. It was clear that the effects were most obvious in the wet parts, hinting that a soil moisture map does a good job differentiating between wet and dry conditions. As hypothesized, elevated planting positions increased seedling survival on wet sites and with decreasing soil moisture content the position became less important, at least for the conifer seedlings. Obviously, low planting positions, like depressions, should be avoided in wet conditions, but with decreasing soil moisture, more flexibility can be applied when planting. In addition, the conifers responded best to the mound position regarding height and diameter. On the contrary, for silver birch it seems like rather the opposite was the key, i.e. avoiding dry positions since mounding did not have a positive effect on neither mortality, height or diameter. Thus, there is a need for more research to understand the effects of site preparation methods on silver birch. The functional ecology of each species has to be considered when developing best practice.
Declaration of openness of research materials, data and code
Data is available upon request from the corresponding author (per.nordin1@gmail.com).
Authors contribution
All three authors contributed to the conceptualization and design of the work. PN did most of the data acquisition and analyzed the data. All three authors participated in the interpretation of data and results. PN constructed the original draft and all three authors participated in reviewing and editing.
Acknowledgements
A special thanks to the FRAS (Future silviculture in Southern Sweden) research program and Mistra Digital Forest for funding P.N to conduct this study. Also, a special thanks to Gisela Björse at Sveaskog and Matts Karlsson at Södra Skogsägarna for assisting with finding the experimental sites. We would also like to thank Erik Olsson for the assistance with field measurements during the spring of 2020.
Funding
This study was made possible through funding from the research programs FRAS (Future Silviculture in Southern Sweden) and Mistra Digital Forest.
References
Ågren A, Lidberg W, Srömgren M, Ogilvie J, Arp P (2014) Evaluating digital terrain indices for soil wetness mapping – a Swedish case study. Hydrol Earth Syst Sci 18: 3623–3634. https://doi.org/10.5194/hess-18-3623-2014.
Ågren A, Lidberg W, Ring E (2015) Mapping temporal dynamics in a forest stream network – implications for riparian forest management. Forests 6, article id 2982. https://doi.org/10.3390/f6092982.
Albrektson A, Elfving B, Lundqvist L, Valinger E (2012) Skogsskötselns grunder och samband. [The forest management series: basics of forest mangement]. Skogsskötselserien 1, Skogsstyrelsen, Jönköping.
Aleksandrowicz-Trzcińska M, Drozdowiski S, Wołczyk Z, Bielak K, Źybura H (2017) Effects of reforestation and site preparation methods on early growth and survival of Scots pine (Pinus sylvestris L.) in south-eastern Poland. Forests 8, article id 421. https://doi.org/10.3390/f8110421.
Bergström R, Bergqvist G (1997) Frequencies and patterns of browsing by large herbivores on conifer seedlings. Scand J For Res 12: 288–294. https://doi.org/10.1080/02827589709355412.
Daugaviete M, Krumina M, Kaposts V, Lazdiņš A (2003) Farmland afforestation: the plantations of birch Betula pendula Roth on different soils. Baltic For 9: 9–21.
European Commission, Joint Research Centre (2022) European atlas of forest tree species. Publications Office of the European Union, Luxembourg. https://doi.org/10.2760/776635.
Fahlvik N, Hannerz M, Högbom L, Jacobsson S, Liziniewicz M, Palm J, Rytter L, Sonesson J, Wallgren M,Weslien JO (2021) Björkens möjligheter i ett framtida klimatanpassat brukande av skog: Sammanställning av nuläget och förslag på insatser för framtiden. [The potential of birch in a future climate adapted forest management: a summary of the presence and suggestions for management in the future]. Report 78, Skogforsk.
Fardusi MJ, Chianucci F, Barbati A (2017) Concept to practice of geospatial – information tools to assist forest management and planning under precision forestry framework: a review. Ann Silvic Res 41: 3–14. https://doi.org/10.12899/asr-1354.
Grossnickle SC (2000) Ecophysiology of northern spruce species: the performance of planted seedlings. NRC Research Press. ISBN 0-660-17959-8.
Grossnickle SC (2012) Why seedlings survive: influence of plant attributes. New Forest 43: 711–738. https://doi.org/10.1007/s11056-012-9336-6.
Grossnickle SC (2016) Restoration silviculture: an ecophysiological perspective: “lessons learned across 40 years. Reforesta 1: 1–36. https://doi.org/10.21750/REFOR.1.02.2.
Grossnickle SC, MacDonald JE (2018) Why seedlings grow: influence of plant attributes. New Forest 49: 1–34. https://doi.org/10.1007/s11056-017-9606-4.
Häggstrom B, Domevscik M, Öhlund J, Nordin A (2021) Survival and growth of Scots pine (Pinus sylvestris) seedlings in north Sweden: effects of planting position and arginine phosphate addition. Scan J For Res 36: 423–433. https://doi.org/10.1080/02827581.2021.1957999.
Hallsby G, Örlander G (2004) A comparison of mounding and inverting to establish Norway spruce on podzolic soils in Sweden. Forestry 77: 107–117. https://doi.org/10.1093/forestry/77.2.107.
Hansson LJ, Ring E, Franko MA, Gärdenäs AI (2018) Soil temperature and water content dynamics after disc trenching a sub-xeric Scots pine clearcut in central Sweden. Geoderma 327: 85–96. https://doi.org/10.1016/j.geoderma.2018.04.023.
Heiskanen J, Saksa T, Luoranen J (2013) Soil preparation method affects outplanting success of Norway spruce container seedlings on till soils susceptible to frost heave. Silva Fenn 47, article id 893. https://doi.org/10.14214/sf.893.
Henneb M, Valeria O, Thiffault N, Fenton NJ, Bergeron Y (2019) Effects of mechanical site preparation on microsite availability and growth of planted black spruce in Canadian paludified forests. Forests 10, article id 670. https://doi.org/10.3390/f10080670.
Hjelm K, Rytter L (2018) The demand of hybrid aspen (Populus tremula× P. tremuloides) on site conditions for a successful establishment on forest land. Silva Fenn 52, article id 10036. https://doi.org/10.14214/sf.10036.
Holmström E, Karlsson M, Nilsson U (2017) Modeling birch seed supply and seedling establishment during forest regeneration. Ecol Model 352: 31–39. https://doi.org/10.1016/j.ecolmodel.2017.02.027.
Holmström E, Gålnander H, Petersson M (2019) Within-site variation in seedling survival in Norway spruce plantations. Forests 10, article id 181. https://doi.org/10.3390/f10020181.
Holopainen M, Vastaranta M, Hyyppä J (2014) Outlook for the next generation’s precision forestry in Finland. Forests 5: 1682–1694. https://doi.org/10.3390/f5071682.
Hynynen J, Niemestö P, Viherä-Aarnio A, Brunner A, Hein S, Velling P (2010) Silviculture of birch (Betula pendula Roth and Betula pubescens Ehrh.) in northern Europe. Forestry 83: 103–119. https://doi.org/10.1093/forestry/cpp035.
Hytönen J, Jylhä P (2005) Effects of competing vegetation and post-planting weed control on the mortality, growth and vole damages to Betula pendula planted on former agricultural land. Silva Fenn 39: 365–380. https://doi.org/10.14214/sf.374.
Hytönen J, Jylhä P (2013) Long-term effect of weed control on survival and growth of silver birch planted on arable land. Baltic For 19: 170–178.
Johansson K, Nilsson U, Allen HL (2007) Interaction between soil scarification and Norway spruce seedling type. New Forest 33: 13–27. https://doi.org/10.1007/s11056-006-9010-y.
Karlsson A (2002) Site preparation of abandoned fields and early establishment of planted small-sized seedlings of silver birch. New Forest 23: 159–175. https://doi.org/10.1023/a:1015605216150.
Karlsson M, Nilsson U (2005) The effects of scarification and shelterwood treatments on naturally regenerated seedlings in southern Sweden. Forest Ecol Manag 205: 183–197. https://doi.org/10.1016/j.foreco.2004.10.046.
Kuglerová L, Ågren A, Jansson R, Laudon H (2014) Towards optimizing riparian buffer zones: Ecological and biogeochemical implications for forest management. Forest Ecol Manag 334: 74–84. https://doi.org/10.1016/j.foreco.2014.08.033.
Langvall O, Nilsson U, Örlander G (2001) Frost damage to planted Norway spruce seedlings – influence of site preparation and seedling type. Forest Ecol Manag 141: 223–235. https://doi.org/10.1016/S0378-1127(00)00331-5.
Lantmäteriet (2022) Lantmäteriet historiska kartor. [Lantmäteriet historical maps], Gävle, Sweden. https://www.lantmateriet.se/sv/kartor/vara-karttjanster/Historiska-kartor/. Accessed 27 June 2023.
Lidman FD, Karlsson M, Lundmark T, Sängstuvall L, Holmström E (2023) Birch establishes anywhere! So, what is there to know about natural regeneration and direct seeding of birch? New For. https://doi.org/10.1007/s11056-023-09971-z.
Larson J, Lidberg W, Ågren AM, Laudon H (2022) Predicting soil moisture conditions across a heterogeneous boreal catchment using terrain indices. Hydrol E Syst Sc 26: 4837–4851. https://doi.org/10.5194/hess-26-4837-2022.
Löf M, Dey DC, Navarro RM, Jacobs D (2012) Mechanical site preparation for forest restoration. New Forest 43: 825–848. https://doi.org/10.1007/s11056-012-9332-x.
Luoranen J, Rikala R, Smolander H (2003) Root egress and field performance of actively growing Betula pendula container seedlings. Scand J Forest Res 18: 133–144. https://doi.org/10.1080/02827580310003704a.
Luoranen J, Viiri H, Sianoja M, Poteri M, Lappi J (2017) Predicting pine weevil risk: effects of site, planting spot and seedling level factors on weevil feeding and mortality of Norway spruce seedlings. Forest Ecol Manag 389: 260–271. https://doi.org/10.1016/j.foreco.2017.01.006.
Luoranen J, Saksa T, Lappi J (2018) Seedling, planting site and weather factors affecting the success of autumn plantings in Norway spruce and Scots pine seedlings. Forest Ecol Manag 419–420: 79–90. https://doi.org/10.1016/j.foreco.2018.03.040.
Månsson J, Kalén C, Kjellander P, Andrén H, Smith H (2007) Quantitative estimates of tree species selectivity by moose (Alces alces) in a forest landscape. Scand J For Res 22: 407–414. https://doi.org/10.1080/02827580701515023.
Margolis HA, Brand DG (1990) An ecophysiological basis for understanding plantation establishment. Can J Forest Res 20: 375–390. https://doi.org/10.1139/x90-056.
Marquis B, Duval P, Bergeron Y, Simard M, Thiffault N, Tremblay F (2021) Height growth stagnation of planted spruce in boreal mixedwoods: importance of landscape, microsite, and growing-season frosts. Forest Ecol Manag 479, article id 118533. https://doi.org/10.1016/j.foreco.2020.118533.
Mc Carthy R, Rytter L, Hjelm K (2017) Effects of soil preparation methods and plant types on the establishment of poplars on forest land. Ann For Sci 74: 1–12. https://doi.org/10.1007/s13595-017-0647-9.
Mohtashami S, Eliasson L, Jansson G, Sonesson J (2017) Influence of soil type, cartographic depth-to-water, road reinforcement and traffic intensity on rut formation in logging operations: a survey study in Sweden. Silva Fenn 51, article id 2018. https://doi.org/10.14214/sf.2018.
Mohtashami S, Eliasson L, Hansson L, Willén E, Thierfelder T, Nordfjell T (2022) Evaluating the effect of DEM resolution on performance of cartographic depth-to-water maps, for planning logging operations. Int J Appl Earth Obs 108, article id 102728. https://doi.org/10.1016/j.jag.2022.102728.
Munson AD, Margolis HA, Brand DG (1993) Intensive silvicultural treatment: impacts on soil fertility and planted conifer response. Soil Sci Soc Am J 57: 246–255. https://doi.org/10.2136/sssaj1993.03615995005700010043x.
Murphy PNC, Ogilvie J, Castonguay M, Zhang C-F, Meng F-R, Arp PA (2008) Improving forest operations planning through high-resolution flow-channel and wet-areas mapping. For Chron 84: 568–574. https://doi.org/10.5558/tfc84568-4.
Nijland W, Coops NC, Macdonald SE, Nielsen SE, Bater CW, White B, Ogilvie J, Stadt J (2015) Remote sensing proxies of productivity and moisture predict forest stand type and recovery rate following experimental harvest. Forest Ecol Manag 357: 239–247. https://doi.org/10.1016/j.foreco.2015.08.027.
Nilsson O, Hjelm K, Nilsson U (2019) Early growth of planted Norway spruce and Scots pine after site preparation in Sweden. Scand J Forest Res 34: 678–688. https://doi.org/10.1080/02827581.2019.1659398.
Nilsson U, Örlander G (1995) Effects of regeneration methods on drought damage to newly planted Norway spruce seedlings. Can J Forest Res 25: 790–802. https://doi.org/10.1139/x95-086.
Nilsson U, Örlander G (1999) Vegetation management on grass-dominated clearcuts planted with Norway spruce in southern Sweden. Can J Forest Res 29: 1015–1026. https://doi.org/10.1139/x99-071.
Nilsson U, Gemmel P, Hällgren J (1996) Competing vegetation effects on initial growth of planted Picea abies. NZ J Forestry Sci 26: 84–98.
Nilsson U, Luoranen J, Kolström T, Örlander G, Puttonen P (2010) Reforestation with planting in northern Europe. Scand J Forest Res 25: 283–294. https://doi.org/10.1080/02827581.2010.498384.
Nohrstedt H (2000) Effects of soil scarification and previous N fertilisation on pools of inorganic N in soil after clear-felling of a Pinus sylvestris (L.) stand. Silva Fenn 34: 195–204. https://doi.org/10.14214/sf.625.
Nordin P, Olofsson E, Hjelm K (2022) Successful spruce regenerations – impact of site preparation and the use of variables from digital elevation models in decision-making? Scand J Forest Res 37: 33–44. https://doi.org/10.1080/02827581.2022.2028895.
Nordlander G, Hellqvist C, Johansson K, Nordenhem H (2011) Regeneration of European boreal forests: effectiveness of measures against seedling mortality caused by the pine weevil Hylobius abietis. Forest Ecol Manag 262: 2354–2363. https://doi.org/10.1016/j.foreco.2011.08.033.
Örlander G, Nilsson U (1999) Effect of reforestation methods on pine weevil (Hylobius abietis L.) damage and seedling survival. Scand J Forest Res 14: 341–354. https://doi.org/10.1080/02827589950152665.
Örlander G, Gemmel P, Hunt J (1990) Site preparation: a Swedish overview. FRDA Report 105, BC Ministry of Forests.
Örlander G, Nilsson U, Nordlander G (1997) Pine weevil abundance on clearcuts of different ages: a 6-year study using pitfall traps. Scand J Forest Res 12: 225–240. https://doi.org/10.1080/02827589709355405.
Pearson M, Saarinen M, Minkkinen K, Silvan N, Laine J (2011) Mounding andscalping prior to reforestation of hydrologically sensitive deep-peated sites:factors behind Scots pine regeneration success. Silva Fenn 45: 647–667. https://doi.org/10.14214/sf.98.
Perälä DA, Alm AA (1990) Regeneration silviculture of birch – a review. Forest Ecol Manag 32: 39–77. https://doi.org/10.1016/0378-1127(90)90105-k.
Persson M, Trubins R, Eriksson LO, Bergh J, Sonesson J, Holmström E (2022) Precision thinning – a comparison of optimal stand-level and pixel-level thinning. Scand J Forest Res 37: 99–108. https://doi.org/10.1080/02827581.2022.2044902.
Petersson M, Örlander G, Nordlander G (2005) Soil features affecting damage to conifer seedlings by the pine weevil Hylobius abietis. Forestry 78: 83–92. https://doi.org/10.1093/forestry/cpi008.
Pikkarainen L, Luoranen J, Peltola H (2021) Early field performance of small-sized silver birch and Scots pine container seedlings at different planting depths. Forests 12, article id 519. https://doi.org/10.3390/f12050519.
Ring E, Ågren A, Bergkvist I, Finér L, Johansson F, Högbom L (2020) A guide to using wet area maps in forestry. Report 1051-2020, Skogforsk.
Rytter L, Lutter R (2020) Early growth of different tree species on agricultural land along a latitudinal transect in Sweden. Forestry 93: 376–388. https://doi.org/10.1093/forestry/cpz064.
Rytter L, Karlsson A, Karlsson M, Stener L-G (2014) Skötsel av Björk, al och asp. [Management of birch, alder and aspen]. Skogstskötselserien 9. Skogsstyrelsen, Jönköping.
Saksa T, Uusitalo J, Lindeman H, Häyrynen E, Kulju S, Huuskonen S (2021) Decision support tool for tree species selection in forest regeneration based on harvester data. Forests 12, article id 1329. https://doi.org/10.3390/f12101329.
Saursaunet M, Mathisen KM, Skarpe C (2018) Effects of increased soil scarification intensity on natural regeneration of Scots pine (Pinus sylvestris L.) and birch (Betula spp. L). Forests 9, article id 262. https://doi.org/10.3390/f9050262.
Schmidt M, Macdonald S, Rothwell R (1996) Impacts of harvesting and mechanical site preparation on soil chemical properties of mixed-wood boreal forest sites in Alberta. Can J Soil Sci 76: 531–540. https://doi.org/10.4141/cjss96-066.
SFA (2022) Swedish Forest Agency geodata, Jönköping, Sweden. https://www.skogsstyrelsen.se/sjalvservice/karttjanster/skogsstyrelsens-geodata/ Accessed 27 June 2023.
Sikström U, Hjelm K, Hanssen KH, Saksa T, Wallertz K (2020a) Influence of mechanical site preparation on regeneration success of planted conifers in clearcuts in Fennoscandia – a review. Silva Fenn 54, article id 10172. https://doi.org/10.14214/sf.10172.
Sikström U, Jansson G, Pettersson F (2020b) Growth responses of Pinus sylvestris and Picea abies after ditch cleaning – a survey in Sweden. Scand J Forest Res 35: 69–84. https://doi.org/10.1080/02827581.2019.1705891.
Simard SW, Jones MD, Durall DM, Hope GD, Stathers RJ, Sorensen NS, Zimonick BJ (2003) Chemical and mechanical site preparation: effects on Pinus contorta growth, physiology, and microsite quality on grassy, steep forest sites in British Columbia. Can J Forest Res 33: 1495–1515. https://doi.org/10.1139/X03-072.
Skovsgaard J P, Vanclay JK (2013) Forest site productivity: a review of spatial and temporal variability in natural site conditions. Forestry 86: 305–315. https://doi.org/10.1093/forestry/cpt010.
Smolander A, Heiskanen J (2007) Soil N and C transformations in two forest clear-cuts during three years after mounding and inverting. Can J Soil Sci 87: 251–258. https://doi.org/10.4141/s06-028.
Sutton R (1993) Mounding site preparation: a review of European and North American experience. New Forest 7: 151–192. https://doi.org/10.1007/BF00034198.
Thiffault N, Jobidon R (2006) How to shift unproductive Kalmia angustifolia – Rhododendron groenlandicum heath to productive conifer plantation. Can J Forest Res 36: 2364–2376. https://doi.org/10.1139/x06-090.
Thiffault N, Titus BD, Moroni MT (2010) Silviculture and planted species interact to influence reforestation success on a Kalmia-dominated site -a 15-year study. Forest Chron 86: 234–242. https://doi.org/10.5558/tfc86234-2.
Thiffault N, Elferjani R, Hébert F, Paré D, Gagné P (2020) Intensive mechanical site preparation to establish short rotation hybrid poplar plantations – a case-study in Québec, Canada. Forests 11, article id 785. https://doi.org/10.3390/f11070785.
Tong CHM, Nilsson MB, Sikström U, Ring E, Drott A, Eklöf K, Futter MN, Peacock M, Segersten J, Peichl M (2022) Initial effects of post-harvest ditch cleaning on greenhouse gas fluxes in a hemiboreal peatland forest. Geoderma 426, article id 116055. https://doi.org/10.1016/j.geoderma.2022.116055.
von Sydow F (1997) Abundance of pine weevils (Hylobius abietis) and damage to conifer seedlings in relation to silvicultural practices. Scand J Forest Res 12: 157–167. https://doi.org/10.1080/02827589709355397.
Wallertz K, Björklund N, Hjelm K, Petersson M, Sundblad L-G (2018) Comparison of different site preparation techniques: quality of planting spots, seedling growth and pine weevil damage. New Forest 49: 705–722. https://doi.org/10.1007/s11056-018-9634-8.
Wiensczyk A, Swift K, Morneault A, Thiffault N, Szuba K, Bell FW (2011) An overview of the efficacy of vegetation management alternatives for conifer regeneration in boreal forests. Forest Chron 87: 175–200. https://doi.org/10.5558/tfc2011-007.
Total of 82 references.
