2.fisiologia II - Neurofisiologia - Parte II
2.fisiologia II - Neurofisiologia - Parte II
Caricato da
FedericoPppzPippoCopyright:
Formati disponibili
2.fisiologia II - Neurofisiologia - Parte II
2.fisiologia II - Neurofisiologia - Parte II
Caricato da
FedericoPppzPippoTitolo originale
Copyright
Formati disponibili
Condividi questo documento
Condividi o incorpora il documento
Hai trovato utile questo documento?
Questo contenuto è inappropriato?
Copyright:
Formati disponibili
2.fisiologia II - Neurofisiologia - Parte II
2.fisiologia II - Neurofisiologia - Parte II
Caricato da
FedericoPppzPippoCopyright:
Formati disponibili
FISIOLOGIA II-VOLUME II
Sommario
FISIOLOGIA II-VOLUME II
Il midollo spinale:............................................................................................................................ 2
Riferimenti anatomici:pag.4-10.................................................................................................. 2
I riflessi spinali:pag.11-22........................................................................................................... 2
Allontanamento dalla noxa:pag. 24................................................................................... 3
Riflesso di Hoffmann:pag.27-28....................................................................................... 3
La locomozione-la postura:pag.35-42,53-62,68......................................................................... 4
Lo shock spinale:pag.66-74......................................................................................................... 5
Il tono posturale:pag.75-85, 92-93, 96....................................................................................... 6
Riassunto delle basi neurofisiologiche del movimento:pag.100-101.......................................... 7
I sistemi piramidale ed extrapiramidale-le mappe:pag.104-131................................................ 7
La sensazione e l'arto fantasma:pag.143-157 ............................................................................ 9
Organizzazione della corteccia motoria primaria: ..................................................................... 9
Le aree motorie secondarie:pag.175 .......................................................................................... 9
I nuclei della base e il talamo:........................................................................................................ 10
I gangli (o nuclei) della base I:pag.43-46.................................................................................. 10
I nuclei della base II:pag.180-211.............................................................................................. 11
Il talamo:pag.184....................................................................................................................... 12
Circuiti della corteccia:pag.183-194.......................................................................................... 12
Patologie associate:................................................................................................................... 13
1.Il morbo di Parkinson:pag.212-216............................................................................... 13
2.I tremori:pag.223-233.................................................................................................. 14
Il sistema limbico; i sistemi della memoria:pag.251-289.............................................................. 16
Riferimenti anatomici:pag.251.................................................................................................. 16
La paura: pag.262...................................................................................................................... 17
Apprendimento e visione:......................................................................................................... 17
La memoria:pag.263-289.......................................................................................................... 18
Tipi di memoria: ............................................................................................................. 18
La localizzazione della memoria:pag.274........................................................................ 19
Il cervelletto:pag.290-348............................................................................................................. 20
Riferimenti anatomici-introduzione:pag.298-300.................................................................. 20
Classificazione:.......................................................................................................................... 21
Struttura fine:............................................................................................................................ 21
Circuiti implicati:...................................................................................................................... 22
Altre funzioni e danni al cervelletto:......................................................................................... 23
Ablazione:pag.323-324............................................................................................................. 24
Esperimenti di stimolazione del cervelletto:pag.325-329........................................................ 24
I sistemi sensoriali:pag.349........................................................................................................... 24
La sensibilità somestesica:pag.353........................................................................................... 25
Il concetto di campo recettoriale:.................................................................................... 25
Le aree somatosensitive:pag.358-362............................................................................. 25
Sensibilità termodolorifica:pag. 367......................................................................................... 25
Il dolore:368.................................................................................................................... 26
Controllo del dolore:........................................................................................................ 27
Oppiacei endogeni e molecole analgesiche:................................................................ 27
Il gate control-la teoria del cancello:pag.375-378...................................................... 27
Controllo discendente:pag.385-389........................................................................... 27
La sensibilità termica:pag.390-394.......................................................................................... 27
I sensi ............................................................................................................................................ 28
L'udito:pag.395-439................................................................................................................. 28
Leggi fisiche e concetti che regolano i suoni:.................................................................. 28
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 1
Riferimenti anatomici:.................................................................................................... 28
Meccanismo di trasmissione:.......................................................................................... 30
Le vie acustiche:410-433................................................................................................. 30
Le zone corticali:pag.414................................................................................................. 30
Udito e linguaggio:........................................................................................................... 31
Le pagine a fianco del nome del capitolo si riferiscono alla dispensa in formato pdf
Il testo in grigio indica delle note aggiuntive, o informazioni utili.
FISIOLOGIA II-VOLUME II
Il midollo spinale:
Riferimenti anatomici:pag.4-10
Pag.18
SN periferico: nervi
SN autonomo: simpatico e parasimpatico che innervano i visceri
SNC: midollo spinale: sistema segmenario (separazione: tentorio) ed encefalo (sistema
sovrasegmentario, dal 12 nervo cranico in su): a sua volta si divide in tronco encefalico (brain
stem) e cervello: costituito da diencefalo e mesencefalo.
SN grigia: è così perchè ci sono le cell ma anche perchè è più vascolarizzata, ecco perchè essume
questa colorazione. E' più vascolarizzata per la metabolismo-dipendenza dell'encefalo (vedi
fisiologia I).
• Sezione a livello di C1: non si può più ventilare perchè c'è sezione tra centri respiratori e
motoneuroni. In + si può avere:tetraplegia:danni a tutti e 4 gli arti (C1). Paraplegia: sezione
midollare a livello toracico, gli arti superiori si muoveranno senza problemi, viceversa per
quelli inferiori.
• Nell'uomo c'è una minor predominanza dell'emisfero sn rispetto alla donna, (il 90% degli
interpreti simultanei è donna).
• Midollo: ha funzioni intrinseche:riflesse, automatiche, di tipo integrativo.
• Regione locomotoria:se in un gatto si asporta diencefalo e telencefalo, lasciando il midollo
spinale, continua a camminare.
• Bell e Magendie: legge di B e M: le radici posteriori sono sensitive e quelle anteriori
motrice-->fu la dimostrazione che i centr nervosi presentano una specializzazione.
• Motoneuroni: quello più piccolo è tonico (per es gamma motoneuroni, nucleo Onuf...):più è
piccolo-->più il sink e la source sono vicini e quindi la soglia è più bassa-->e quindi maggior
eccitabilità.
Soglia bassa, maggior eccitabilità, soglia alta: minor eccitabilità. Il principio delle
dimensioni, unito al concetto di sink-source, fa capire il fenomeno del reclutamento di fibre,
un fenomeno di graduale aumento di forze. Più il neurone è piccolo, e più la sink-source è
vicina, e viceversa (il cosidetto size principle).
02.12
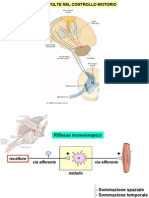
I riflessi spinali:pag.11-22
Grazie ai riflessi si può studiare la funzionalità del SNC, soprattutto del midollo.
Si ricorda la strutturazione del midollo in lamine di Rexed, ossia precise stratificazioni di neuroni.
In ogni caso le corna posteriori sono deputate a ricevere segnali in ingresso e viceversa quelle
anteriori.
Il riflesso spinale è un riflesso in cui si ha una via afferente al midollo, un centro di elaborazione e
una via afferente. Il riflesso per definizione deve essere innato ed involontario.
Classificazione:
• Riflesso monosinaptico: sinapsi unica tra le vie (riflesso da stiramento miotatico
antigravitario, legato ad attività dei fusi neuromuscolari). Prevede un'unica sinapsi tra
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 2
neurone di ingresso e di uscita. Il primo neurone è nel ganglio spinale (cell a T): avviene
attraversamento poi di corteccia cerebrale e cerebellare. (Ricorda le vie spinocerebellari, il
nucleo di Clarke per es, oppure area 3A della corteccia cerebrale). Le afferenze dei fusi
inviano fibre ascendenti (colonne posteriori) a tali zone. Non avviene nei motoneuroni una
scarica maggiore di 150 impulsi/s per la presenza di interneuroni di Renshawn: sono
interneuroni inibitori, che secernono glicina. Si genera un feedback negativo, perchè la loro
attività è dir prop all'attività di scarica del motoneurone.
Lo stimolo di stiramento arriva al midollo spinale, e in via riflessa, ha 5 possibilità: per es le
afferenze dei fusi possono finire (a) al cervelletto, e con i fasci lemniscali alla corteccia (b),
oppure (c) ai muscoli, che devono essere mantenuti a lunghezza costante: qualunque forza
si riflette sul muscolo omonimo e sui muscoli sinergici (riflessi monosinaptici, che
provocano contrazione), e poi a un motoneurone del muscolo antagonista, che deve essere
però inibito. Una contemporanea contrazione di flessori ed estensori è sintomatico di
patologie (per es a gangli della base).
• Riflesso polisinaptico: di tipo antinociocettivo soprattutto, ossia per allontanarsi da una
fonte di dolore. (vedi sotto, allontanamento da noxa).
• Riflesso patellare:pag.22: la percussione va fatta sul tendine del quadricipite, sotto la rotula,
eseguendo manovra di Jendrassik, si può testare la funzionalità midollare-->infatti, si avrà
movimento della gamba non voluto dal paziente. E' possibile fare un analogo test a livello
del tendine di Achille (si stirano i fusi muscolari del gastrocnemio, e si avrà estensione verso
l'esterno del piede).
• Se si colpisce il processo mentoniero si avrà il riflesso mentoniero (per testare V nervo
cranico).
• Riflesso di grattamento: già detto, vedi indietro.
• Riflessi appresi: necessaria la conoscenza pregressa dell'evento, sono secondari.
• Alfa motoneuroni: grandi, con ramificazioni di 300-500 micron. Ricorda il concetto della
via finale comune, tutte le vie discendenti che coinfluiscono, per dare l'atto, tramite i
motoneuroni alfa. (mentre i gamma sono responsabili del tono).
Allontanamento dalla noxa:pag. 24
esiste un gruppo di riflessi la cui funzione è quella di sfuggire alle noxe, ossia a fattori nocivi o di
danno: nel riflesso antinociocettivo i recettori attivi sono termici e dolorifici (fibre di III-IV gruppo
di Lloyd). L'allontanamento è dato dalla stimolazione dei recettori. Il dolore può essere dovuto o a
stimolazione di recettori termici o dolorifici o a iperstimolazione di recettori tattili.
La noxa attiva quindi motoneuroni flessori, viceversa verranno inibiti gli estensori, tramite un
meccanismo però polisinaptico. Nell'altro lato del corpo si avrà, viceversa, un riflesso estensore
crociato, per es se trovo per terra un chiodo e devo evitarlo. In tal modo si fornisce al corpo un
piano di appoggio, e si evita caduta (per principio di doppia innervazione reciproca).
Dato che è polisinaptico, siamo di fronte a fenomeni di tipo divergente, per es vengono emessi urli
di fronte a un dolore (grazie al PAG, periaqueductal grey): l'urlo è un segno di allarme, di richiesta
di aiuto verso gli altri.
Pag.26
Riflesso di Hoffmann:pag.27-28
Schema di Hoffman: serve per capire il tempo del riflesso, misurandolo in millisecondi. Tra stimolo
e risposta ci sono 30 millisecondi: vengono appoggiati 2 ellettrodi (150 microampere), in modo tale
da stimolare le fibre nervose sottostanti (stimolazione transcutanea), a livello dell'arto inferiore per
esempio. Le fibre di un nervo come detto hanno diametro differente:quando si raggiungono i 150
microampere, per 1 millisecondo, avverrà la stimolazione delle fibre più grosse. Si dice che è
liminare per eccitare le fibre di diametro più grosso, che sono a maggior velocità di conduzione e a
soglia più bassa.
Le fibre più grosse sono quelle dei fusi neuromuscolari (20 micron), che verranno così stimolate (è
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 3
come se avessi colpito il tendine quindi).
Si attiveranno poi i motoneuroni connessi con i fusi, e poi scatta l'arco riflesso che verrà registrato
come un'onda di depolarizzazione dagli elettrodi stimolanti (che avranno così una doppia
funzione).
Si avrà una rappresentazione su carta millimetrata, grazie alla quale si può calcolare il tempo che
trascorre tra la stimolazione e l'effetto.
Aumentando l'amperaggio si ecciteranno altri tipi di fibre (per es: 200 microA: fibre di alfa
motoneuroni).
In questo ultimo caso avrò eccitazione del muscolo diretta, grazie agli alfamotoneuroni, e
indirettamente da arcoriflesso H. La risposta diretta sarà anche in anticipo rispetto ad H.
In più l'arcoriflesso H diminuisce di intensità: infatti, stimolo le fibre portatrici dello stimolo del
fuso neuromuscolare, nonchè fibre che portano lo stimolo in 2 vie, in avanti e in via antidromica.
Questo ultimo porta a uno "scontro" tra un evento ortodromico e uno antidromico, si avrà quindi
un blocco della via riflessa, del riflesso tramite il sistema miotatico (fenomeno della collisione tra
riflesso ortodromico miotatico e quello antidromico dello stimolo riflesso-legge della conduzione
indifferente).
Maggiore sarà l'intensità dello stimolo, minore sarà la risposta H quindi; a 300 microampere si
blocca totalmente la conduzione: la collisione tra la stimolazione cellulipeta o antidromica azzera
l'attività ortodromica, normale e fisiologica del neurone (la risposta H). Questo si verifica in
condizioni sperimentali; si comprende così come l'assone non sia solo unidirezionale: lo stimolo si
propagherà per via cellulifuga e cellulipeta.
Test della stimolazione antidromica: utile spia per studiare eccitabilità dei neuroni.
• pag.97-99:Soggetto su piattaforma mobile: si studia la reazione per stabilizzare la
posizione: il punto di stabilità è la caviglia, nonchè il bacino, che è un altro punto di
controllo. Tutti questi movimenti dipendono in ogni caso dai transcortical "reflex", (non è
propriamente un riflesso, perchè non è presente fin dalla nascita). Riflesso di Hoffman: 30
millisecondi, tale transcortical reflex dura 100 msec.
• Il riflesso da stiramento permette di stare in piedi: quando siamo sui talloni però, è
necessario spostare il baricentro.
La facilitazione e la occlusione:
la facilitazione dello stimolo dovuto alla sommazione delle vescicole esocitate. E' correlato ai
fenomeni di sommazione spaziale e temporale, grazie ai quali 2 o più stimoli subliminali da soli
riescono a generare insieme uno spike.
L'occlusione, ossia fenomeno opposto al primo, in cui il neurone non riesce a rispondere a degli
stimoli eccessivamente vicini e contrari. E' l'occlusione degli impulsi.
03.12
La locomozione-la postura:pag.35-42,53-62,68
C'è sempre una spinta in avanti. Il riflesso del gastrocnemio ci aiuta nel movimento.
Nei pazienti cerebellari o con ictus, si fanno delle prove verso un muro, verso cui si spinge il
paziente, in modo tale che non caschi (perchè non avrà il riflesso detto transcortical loop: è un
riflesso appreso in infanzia (3-4 anno), che viene perso o rallentato in caso di pazienti affetti da
parkinson o cerebellari, o ai gangli della base:sono dei riflessi che però implicano corteccia
cerebrale, che permette la stabilizzazione.
Esperimento su gatto: si può regolare velocità del tappeto sotto le zampe, per vedere la diversa
reazione che ha.
Si nota che c'è una regione locomotoria, situata nel mesencefalo (questo perchè la zona locomotoria
è vicina ai collicoli inferiori, stazioni acustiche-->scatta infatti un fenomeno di locomozione
correlato alla difesa, che si mette in moto grazie a delle percezioni acustiche; in più c'è una
correlazione con ipotalamo posteriore (corpi mammillari del circuito di Papez) zone, in animali
inferiori, deputate a funzioni sessuali-->si attivano le vie di movimento, l'animale sarà diretto verso
la compagna da conquistare). In generale nel mesencefalo risiedono strutture che permettono il
"go", l'azione).
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 4
Si fa un'altra sezione, a livello spinale. Si è separato rigonfiamento lombo sacrale dalle zone
superiori: l'animale, non si muoverebbe in teoria, ma, se si stimola l'estremità delle zampe, queste
iniziano a muoversi.
Allo stesso tempo si registra l'attività tramite elettromiogramma: si ha attività elettrica e quindi
meccanica, come al solito. Il gatto sta al passo, si muove con calma, la fase di oscillazione è più
breve della fase di appoggio; se però si modifica la velocità come detto in precedenza, si noterà
come la fase di flessione, di oscillazione,m è costante, indipendentemente dal contesto. Varia la
parte nera, ossia la posa a terra. Questo perchè la componente flessoria ha una forte dipendenza del
SNC, (concetto di programma centrale). Anche la prima parte della deglutizione dipende da un
programma motorio, a livello esofageo.
Quindi il movimento è caratterizzato da una parte di azioni stereotipate, fisse, meccaniche, e altre
dipendenti dalla volontà, dalla coscienza.
Pag.42
• Central pattern generators: (CPG),pag.40: sono circuiti oscillanti, devono oscillare
tra 2 azioni, come per es masticazione, il passo, la respirazione...Sono avviati dalla regione
locomotoria, per quanto riguarda il movimento. Ricevono impulsi da zone superiori, senza
di cui ci sarebbe un andamento casuale, erratico.
• Deambulazione nel bambino:3 ipotesi di sviluppo: nel bambino, più si cresce e pian
piano più facile sarà deambulare, grazie a riduzione delle dimensioni della testa (ipotesi
fisica). Da un altro lato (ipotesi cognitiva) si è visto che c'è un rapporto inverso tra capacità
motoria e cognitiva, più veloce il primo, e più lento è il 2. Infine c'è una ipotesi
neurofisiologica: la mielinizzazione non è completa nel bambino (ecco perchè si avranno i
riflessi visti in precedenza); in più nel bambino i centri superiori sembrano inibiti, ecco
perchè non cammina da se. Infine, nel bambino c'è una maturazione craniocaudale, è più
importante sviluppare la funzione masticatoria che quella deambulatoria.
09.12
Lo shock spinale:pag.66-74
Così chiamato perchè si ha paralisi flaccida, con perdita del tono, oppure areflessia, perchè manca
tono di base dei neuroni.
Il cervelletto è una importante struttura tonigena nei confronti di diverse strutture.
In più si potrà avere anche un calo di pressione arteriosa media a causa di tale shock.
Compaiono i riflessi patologici come quello di Babinski, vescica va incontro ad una evacquazione
riflessa, idem per l'ampolla rettale. Scompare l'erezione psicogena resta quella riflessa.
• Il riflesso di Schiff-Sherrington:nei quadrupedi i segmenti lombari hanno effetto inibente
su quelli toracici: ipertonia degli estensori degli arti superiori, e paraplegia che risulta da
lesioni compressive, acute del midollo spinale, causando rimozione delgli effetti inibitori
dei neuroni del tratto lombare.
• Opistotono:rigidità dei muscoli nucali, iperflessione dorsale, importanti per non far
"cadere" testa in basso (negli animali, in cui la testa è dietro): si vede anche negli umani,
ossia è uno spasmo momentaneo che colpisce i muscoli estensori del dorso che comporta
l'iperestensione del capo e talvolta dell'intera colonna sull'individuo supino.Si verifica
durante le meningiti, il tetano, la tetania e anche nell'isteria. (da internet).
• Sindrome di Brown-Sequard:emisezione: rara, per es se un proiettile attraversa il midollo
tangenzialmente, non in centro, quando lo sfiora: consiste in paralisi flacida dell'emisoma
ipsilateralmente, anestesia somestetica lemniscale, termodolorifica dell'emisoma
controlaterale (caudalmente alla emisezione spinale scompaiono dallo stesso lato quelle
funzioni che dipendono da sistemi che si incrociano al di sopra della sezione ossia il fascio
corticospinale e il sistema lemniscale, mentre dal lato opposto scompaiono quelle funzioni
che dipendono da sistemi che si incrociano sotto la sezione (sistema anterolaterale)).
• Rigenereazione del midollo spinale:pag.71-74: la microglia cicatrizza l'assone e blocca la
ferita. E' sempre la microglia che produce no go factors, che inibiscono l'accrescimento
assonico. Fondamentale è la plasticità neuronale in tutto ciò ,se i neuroni hanno un target
riescono a raggiungerlo. Bisogna però fermare i NO GO factors, che impediscono ciò.
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 5
http://comps.fotosearch.com/comp/LIF/LIF119/brown-sequard-syndroom_~BRNSQRD.jpg
http://www.aan.com/familypractice/html/Chapter1%20The%20Neurologic
%20Examination_files/image032.jpg
http://medinfo.ufl.edu/year2/neuro/review/images/fig02.jpg
http://www.univ-
brest.fr/S_Commun/Biblio/ANATOMIE/Web_anat/Snc/Moelle_Epi/Syndro_Brown_Sequard.j
pg
Il tono posturale:pag.75-85, 92-93, 96
E' fondamentale per contrastare la gravità, è mantenuto dai muscoli: 2 nuclei di riferimento :
sistema reticolo spinale e i nuclei reticolari bulbari e pontini e il nucleo vestibolare di Dayters (vie
reticolospinali e reticolobulbari): Deiters è una struttura bulbare, la reticolare è
bulbopontomesencefalica. Il controllo finale del tono spetta a tutte queste strutture, con una
reglazione gamma (reticolare), alfa (vestibolare).
Sherrington per primo ha studiato ciò, tramite animali decerebrati (sezione intercollicolare del
mesencefalo): si è visto che aveva ipertonia di tutti gli arti, come se si avesse liberato i centri del
tono posturale (strutture pontine e bulbari) da strutture superiori inibitorie, i gangli della base. E'
una release dei sistemi antigravitari: nell'uomo si avrà una posizione con arti superiori flessi. (No
inibizione su reticolare facilitante, no eccitazione su reticolare inibente. )
Arrivato a livello del 12 nervo cranico, si avrà lo shock spinale-->da questi 2 tagli si è compreso
dove fossero localizzate le strutture deputate al controllo posturale.
E' implicata pure la formazione reticolare, che ha 3 infulenze: 1. discendente, con vie reticolo
spinali, soprattutto gamma, 2. influenza ascendente sui sisitemi della veglia, dell'arahusal...
Reticolare mesencefalica-pontina: risveglio, è attivante
Reticolare bulbare (zone di Jouvè(?) :inibenti, ricche di serotonina. Nucleo di Magoun-Rhines:
serotoninergico, se stimolato scatena il sonno-->inibizione gamma motoneuroni.
3.Infine ha controllo del dolore, ha effetto sui neuroni encefalinergici.
• Nucleo rosso: se viene lasciato nel sistema, non si verifica la rigidità, e l'arto ha una
consistenza "cerea", non cede improvvisamente: se si seziona al di sopra di tale nucleo, esso
è attivo: si proietta ai motoneuroni di tipo flessore-->se si piega arto si avrà azione sia dei
flessori sia degli estensori (questo nel gatto, in cui il nucleo rosso proietta ai flessori nel
gatto). Si avrà, insomma, una tensione ipertonica di tutti i muscoli, gravitari e antigravitari.
Nell'uomo tale nucleo ha una diversa funzione, diverso target (questo per plasticità
filogenetica, fenomeno simile al fatto che l'ippocampo nelgli animali è legato ad attività
sessuale, mentre nell'uomo è importante per valutare memorizzazione di fenomeni
emozionalmente coinvolgenti). Infatti vengono favorite così le vie rubrolivocerebellari e
viceversa, chiudendo il circuito a feedback, dal cervelletto al nucleo rosso.
Il controllo dei flessori, durante il movimento, è affidato al fascio piramidale, e non al
nucleo rosso: ciò sarà visibile soprattutto in una lesione del fascio extrapiramidale (con
implicazione anche dell'area premotoria, del giro pre e postcentrale appartenenti al sistema
estrapiramidale, insieme ai nuclei della base). Questi elementi di tale fascio scaricano sullo
striato, formando il sistema di movimento dei gangli della base e del sistema SEPOC.
• Riflesso miotatico inverso o di Philiphson (o del coltello a serramanico): arto esteso a
rischio di strappo, stiramento del muscolo o del tendine-->tramite interneuroni inibitori si
riduce la contrazione. La via afferente è l'organo del Golgi. E' formato da 3 neuroni, di cui
uno inibitore. Il cedimento è improvviso. Lo stiramento eccessivo del muscolo invece che
portare a contrazione porta a una decontrazione, o caduta di stato contrattile, di natura
protettiva, per evitare rottura del muscolo teso o lo strappo del tendine correlato.
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 6
Pag.90
• Se un animale fosse in uno stato di rigidità, è sufficiente lesionare radici posteriori
(sensitive)per capire se c'è una rigidità gamma:scompare sezionando tali radici (anello
gamma alfa si chiude in tal modo). Se invece la rigidità resta anche in questo caso, non sarà
una rigidità gamma. Ovviamente se sezione radici ventrali, scomparirà in ogni caso.
15.12
Riassunto delle basi neurofisiologiche del movimento:pag.100-101
I:
Atti motori riflessi, innati, per es la suzione, nel bambino.
Atti motori volontari: hanno un controllo soprasegmentario continuo
Atti motori automatici: origine volontaria, sono sequenziali, modificati dall'apprendimento
(guidare per es, dopo aver imparato non è più necessario uno stretto e attento controllo).
II:
Attività motorie di tipo tonico:postura.
Attività motorie di tipo fasico: movimento
III:
Motililità assiale: attività tonica di tipo posturale: stare in piedi, stazione eretta in deambulazione
Motilità prossimale: attività tonica e fasica, scelta di aggiustamento posturale adeguato (implicato
paleostriato, FR, nucleo rosso...)
Motilità distale: scrittura, attività motorie fini (implica vie piramidali, neostriato, cervelletto).
I sistemi piramidale ed extrapiramidale-le mappe:pag.104-131
I sistemi extrapiramidali sono formati da numerose stazioni.
E' detto piramidale: perchè si incrocia a livello delle piramidi bulbari, solo per il 10% è diretto, resta
anteriore, non ha incrocio.
Nell'uomo e nei primati le vie discendenti si dirigono ai nuclei motori dei nervi cranici.
Il fascio piramidale è formato da 5 milioni di fibre, solo un 1/5 diventa fascio corticospinale. 4
milioni restano quindi a livello cranico.
Deputata alla fine regolazione motoria e alle afferenze dirette alla corteccia.
Le fibre provengono per il :60%: da area 4 gamma e area prerolandica, (circonv. Precentrale). 20%
area 6, e per il 20% da area 2,3 (vie sensitive). Quindi da aree rolandiche e perirolandiche, intorno
alla scissura di Rolando. Nel fascio piramidale ci sono neuroni piccoli, tonici, facilmente eccitabili,
sono anche tonigeni quindi, danno un tono di base.
La zona anteriore è deputata agli output, è detta sintagmatica, è motrice,l'altra, posteriore,
paradigmatica, deputata a ricevere gli input. I neuroni sono disposti secondo un ordine preciso,
viene tenuto conto del cosidetto principio dell'economia, "della via più breve possibile". Le zone
della corteccia che controllano le dita e la bocca, sono prossimiori, vicini, per il fatto che spesso i
movimenti di queste due parti del corpo avvengono in simultanea.
Si sono ricostruite diverse mappe neuronali, (vedi per es homunculus sensitivo e motorio): tutti i
sensi con relay talamici hanno una organizzazione corticale somatotopica (per tutti tranne per
l'olfatto). Le mappe hanno una estensione funzionale, più una zona è funzionalmente importante, e
più sarà estesa nella mappa. Faccia e mani: sono per es più del 50% della mappa corticale.
E' da notare che le mappe si sono create solo negli animali allattanti, che allattano, e che hanno
quindi dei rapporti interpersonali, se c'è dell'intersoggettività. (uccelli e anfibi per es non avranno
tali mappe). Da notare la disposizione particolare del corpo nella rappresentazione (corpo
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 7
capovolto e controlaterale...dovuto alle connessioni talamo corticali): questo perchè nel talamo si
ha incrocio dei lemnischi per es (o gracile, cuneato...)... le fibre si incrociano: le porzioni
toracocervicali, che erano laterali, diventano mediali.
In più si avrà rotazione delle vie talamocorticali, le fibre della faccia vanno verso il basso e viceversa
quelle delle parti più caudali del corpo (sacrali). Se così non fosse le fibre del volto non uscirebbero
dal talamo.Di conseguenza anche il talamo avrà una somatotopia.
Sembra che ci siano delle sostanze che attraggano le fibre, con una certa specificità: sono delle
molecole segnale, che dal talamo portano le fibre verso la corteccia.
Le mappe sono multiple, come detto: esistono anche nel cervelletto, è capovolta per quanto
riguarda il verme ma non è capovolta per i lobuli paramediani. Esistono anche quelle
retinotopiche...e altre, sono comunque diverse. Le prime mappe sono state costruite stimolando le
varie parti del corpo (Penfield). Tale molteplicità è fondamentale nella riabilitazione per es.
Mentre l'homunculus sensitivus è stato costruito stimolando le parti della corteccia, nelle apposite
aree (la corteccia proiettava impulsi alla parte del corpo interessata).
A questo si ricollega per es la sindrome dell'arto fantasma...
Nella scimmia si vede un'organizzazione anulare, ad imbuto.
Nell'uomo si è studiato tutto tramite la marcia Jacksoniana dell'epilessia (movimento da dita a
gomito).
Durante la neurogenesi, come è possibile che le fibre si connettano correttamente tra di loro?
Tramite spiecifiche molecole proteiche. Vedi più avanti.
• Visione: tramite stratificazione successiva di impulsi nervosi, che passando da un area
all'altra si arricchisce sempre più di informazioni.
• Vibrisse nel topo:tatto attivo. Le vibrisse sono degli organi sensitivi particolari, sensibili alle
onde pressorie di un eventuale aggressore. Per studiarle si sono usati aminoacidi marcati
nelle vibrisse-->vengono portati nel nucleo trigeminale, nel gracile, cuneato... In più nei
ratti piccoli sono state rimosse le vibrisse. Poi si sono messi aa radioattivi nei follicoli-->i
"barrels" vicini si sono ingrossati, grazie ad apposite neuromolecole che hanno causato lo
sprouting dei neuroni vicini. C'è stata una colonizzazione da parte delle cell vicine alla zona
in cui ho operato l'asportazione.
• Le aree del cervello si modificano in base alla stimolazione (dai una letta a pag. 127).
• In un altro esperimento si è sezionato il nervo mediano, non ci saranno ovviamente più
afferenze a livello corticale. Dove non c'è più la risposta, ci sarà un'occupazione della zona
da parte del nervo radiale. La degenerazione stimola le altre zone tramite apposite sostanze
eccitanti i neuroni vicini, che vanno a ricolonizzare le zone.
• La riabilitazione può quindi portare ad un nuovo trofismo, ad una rigenereazione, se si
stimolano altre zone prossimali a quelle coinvolte dal danno. Però, la riattivazione di zone
mancanti farà si che ci sia una proiezione in zone che una volta erano presenti (vedi
discorso dell'arto fantasma: i neuroni che erano dell'arto, liberano neurotrofine e altri GF--
>i neuroni vicini a quelli dell'arto proliferano (per es quelli del volto)-->ora, se si stimola
per es il mento del paziente, egli percepirà per es il pollice. Si ha attivato le zone faccia, ma
per "rimbalzo" si ha attivato zone e neuroni che erano delle parti del corpo amputate. E' una
attivazione per "interferenza" si può dire (cfr sezione oltre).
• Solco centrale: divide il cervello in 2 parti:1.Efferente, sintagmatica, 2.Afferente,
paradigmatica.Da qui si può comprendere dove sono presenti lesioni: problemi motori--
>zona 1 ; problemi di agnosie-->zona 2
16.12
• La funzione di tale via è esplicata durante i movimenti di precisione.
• L'uomo, con le sue mani, è capace sia di afferrare (grasping), sia di opporre il pollice: in
particolare quest'ultima abilità è tipica solo dell'uomo.
• Il fascio piramidale garantisce anche la secrezione (minima) di sudore, per esempio a livello
delle dita, della mano intera, oppure dei piedi e del volto. Il resto del corpo è ovviamente
controllato dai centri termoregolatori.
• Nell'evoluzione è aumentato in dimensioni, e viceversa, si sono ridotte le vie rubrospinali;
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 8
inoltre, nell'evoluzione si sposta dal midollo al cervelletto.
• Tale fascio, in senso craniocaudale, si ramifica. Nell'encefalo, si liberano collaterali ai gangli
della base, all'amigdala, al talamo, ai nuclei gracile e cuneato, ai collicoli superiori, al nucleo
rosso e al cervelletto (pag.164).
• Sono vie dette corticipete, ma sono controllate dal fascio piramidale, che è una sorta di filtro
per tutto ciò che raggiunge l'encefalo. Si origina dalle zone prerolandiche, e controlla ciò che
afferisce a tali strutture.
• Il glutammato e i recettori ampa sono gli elementi più utilizzati in questo sistema.
La sensazione e l'arto fantasma:pag.143-157
Per es la percezione di una puntura di spillo: viene percepito il dolore dopo 20-25 ms, perchè lo
stimolo deve arrivare alla corteccia e generare la sensazione. Il cervello ovviamente deve
interpretare tale stimolo proveniente dall'esterno, e ci permette la percezione.
I recettori avranno quindi un doppio meccanismo di funzionamento: possono o "subire", una
percezione, essere passivi, oppure essere attivi, nel caso in cui ci sia una azione funzionale alla
percezione, quando il soggetto esegue attivamente un atto (come nel caso del braille e dei ciechi).
L'arto fantasma è quel fenomeno rilevato in pazienti sottoposti ad amputazione di un arto.
I neuroni associati alla mano non avranno ovviamente più stimoli; tali cellule producono GF, che
stimolano i neuroni presenti nella zona, per es quelli del volto: questi ultimi si avvicineranno ai
neuroni della mano: toccando il volto del paziente, è possibile che esso rilevi una sensazione come
se provenisse dalla mano.
Tali eventi si manifestano per 10 anni circa, dopo i neuroni muoiono e non si verificherà più tale
fenomeno.
• Sherringhton compì degli esperimenti di stimolazione del fascio piramidale sempre tramite
stimoli elettrici.
• Stimolazione di aree attorno alla zona da operare: viene eseguita per capire che aree sono
presenti nella zona, e per evitare di danneggiarne alcune fondamentali. Interessante notare
come una stimolazione di tale tipo non permette la ripetizione dell'atto, cosa che
ovviamente avviene in natura, fisiologicamente: si parla di instabilità dei punti corticali,
nonchè dell'importanza della volontà.
• Il movimento: la corteccia coordina i muscoli in modo tale da creare un quadro completo
della situazione in cui ci si trova. Si può ragionare in termini di modello a mosaico.
• Se viene lesa la via piramidale causa lentezza, perdita di forza nel movimento, perdita dei
movimenti fini.
• Tono piramidale: generato soprattutto dall'azione tonica e facilitante del cervelletto, sulla
corteccia cerebellare.
• Il fascio piramidale è, come anticipato, capace di controllare gli stimoli provenienti da
un'area: si attivano i neuroni a livello dei nuclei di Goll e Burdack e da qui lo stimolo viene
controllato dal fascio piramidale.
• La via piramidale attiva quella reticolare, che a sua volta attiva la corteccia, si forma una
sorta di circolo di riattivazione.
• I neuroni motori sono "consapevoli" dell'azione che eseguono e del risultato: ci sarà cioè un
collegamento tra l'azione esterna e quella interna: ossia c'è una connessione tra la zona
prerolandica e quella postrolandica--> l'individuo è cosciente così dell'azione, per es la
presa di un oggetto.
Organizzazione della corteccia motoria primaria:
gli stimoli efferenti si dipartono dal 5 strato della corteccia cerebrale.
Una colonna corticale attiva un micromovimento, e tante colonne attivano tante piccole porzioni di
tanti muscoli-->da qui si scatenerà un movimento complesso.
Le aree motorie secondarie:pag.175
Area 6: premotoria, fondamentale per la programmazione delle sequenze motorie: non si attiva nel
caso del movimento semplice, mentre in altri atti più complessi si attiva (contare con le dita).
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 9
Anche il solo pensiero del movimento farà attivare l'area 6.
Tali azioni programmate sono vicine nel tempo: vedi per es nel caso dei movimenti saccadici
dell'occhio, a cui segue l'attivazione dei campi oculomotori frontali che permettono la messa a
fuoco dell'oggetto, e poi il movimento del corpo per avvicinarsi all'oggetto.
A questo punto si dovrà verificare anche l'arresto sul target, che sarà dovuto all'attivazione dei
muscoli antagonisti (regolazione della sinergia muscolare).
Lesioni corticali:pag.158
ablazione corticale localizzata, per es all'area 4, causa, nei primati:
1.flaccidità degli arti
2.scomparsa controlaterale di riflessi mono e polisinaptici
3.Comparsa controlaterale di riflessi patologici, come segno di Babinski...
Si avrà sia perdita del movimento (paresi), sia del tono (flaccidità, disfacilitante il movimento.
Lesione del fascio corticospinale:
Dopo alcuni giorni dalla lesione si verifica mancanza di movimenti fini, bradicinesia, mancata o
ridotta forza, nonchè ipertonia selettiva, compare la cosidetta ipertonia selettiva.
17.12
• Pag.177-179:Aree 6, 5, 7 (area motoria supplementare, importanti per il programma
motorio). La massima attività in tali aree è raggiunta durante la programmazione del
movimento, non quando è già avvenuto.
• Riassumendo (immagine sotto):
• La corteccia passa il comando ai motoneuroni "eseguendo" il piano motorio. Il cervelletto è
fondamentale per eseguire il movimento. (cervelletto, gangli base: sono extrapiramidali,
intervengono nella fase iniziale (cervelletto, ma esiste anche il lateal cerebellum, implicato
nella programmazione), nella pianificazione o nell'esecuzione (gangli)).
• Pag.195-196:ree e campi personale/extrapersonale: area 6 importante per controllo di
movimenti in area personale, (a seguito di lesione si verifica grasping reflex) l'8 per quelli
extra (FEF frontal eye fields).
Pag.178
Pag.179
I nuclei della base e il talamo:
I gangli (o nuclei) della base I:pag.43-46
Caudato+putamen=striato (così detto perchè ha gli striosomi, ossia le zone influenzate dal sistema
limbico).
Non sono solo un gruppo di controllo di movimento, ma hanno 4 funzioni:
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 10
1.sensitivo motorio
2.emozionale limbico
3.cognitivo
4.oculomotorio
Sono fondamentali nella programmazione e organizzazione del movimento, (mentre cervelletto è
in azione durante e dopo il movimento, in ogni caso a movimento iniziato, la sua posizione è
fondamentale per la sua funzione).
Striato: riceve anche da centrum medianum ,la corteccia cerebrale ha connessioni dirette con esso.
Nel grasp reflex del neonato, si ha un riflesso di natura extrapiramidale (area 6, come detto,
SEPOC, vedi I modulo fis II). Tutti i gangli della base appartengono al sistema
extrapiramidale(controlla azioni posturo-cinetiche), ossia fuori dal fascio piramidale (che controlla
fine motilità distale).
Il parkinsoniano ha una tipica andatura oscillante, inizia e termina con lentezza una azione di
movimento.
Sono caratterizzati da un sistema di doppia inibizione, ossia (-)(-)-->+ , si avrà una disinibizione,
una attivazione praticamente. Si ha un sistema GABA con GABA, una inibizione talamica, che a sua
volta viene inibito dal caudato-putamen. In questo modo si risparmia sulle vie, si regola solo il
freno, non è necessario aggiungere "l'acceleratore".
Dopamina: attiva il sistema striato, per il movimento, mentre, agendo su accumbens/lobo frontale,
attiva zone prefrontali e sopraottiche. Nucleo accumbens: via dopaminergica spinolimbica
(dopamina implicata negli animali anche a funzioni sessuali).
Pag.46
• Nuclei VA VL del talamo: V sta per ventral, sono nuclei specifici, che si proiettano cioè ad
aree specifiche della corteccia. VA è più frontale (quindi connesso ad aree di tipo
associativo), mentre VL è motrice.
• Sistema gamma: è un sistema che può fungere anche da preattivatore degli alfa (anello
gamma alfa, vedi I modulo).
• Uomo in coma: uomo che non ha più coscienza, non sente più il dolore.
I nuclei della base II:pag.180-211
Fondamentali nel SEPOC (il sistema extrapiramidale di origine corticale, comprende molte zone 8
6 4s, 1 2 3, 7a e 7b; è composto da numerose stazioni: è polisinaptico, parte dalla corteccia
cerebrale.Le fibre che si originano da tali zone convergono verso il sistema neostriatale, verso il
caudato e putamen, quindi verso i gangli della base.
Deputati insieme al cervelletto all'elaborazione del programma motorio.
Da qui si formano fibre che saranno dirette verso i fasci rubrospinali, nigrostriatali, reticolospinali,
che si portano poi ai motoneuroni alfa e gamma.
I nuclei, come anticipato, hanno funzioni inibenti nei confronti del movimento: la rigidità da
decerebrazione è una spasticità, per liberazione delle vie gamma motorie.
I nuclei servono per organizzare il movimento, per pianificarlo dal punto di vista fisico.
Le afferenze visive sono fondamentali per il movimento-->esiste una via dorsale dell'area occipitale
che proietterà al SEPOC: l'area 6 e il SEPOC servono per trasformare afferenze visive in una
efferenza motoria. (vedi il caso in cui si è al buio e ci si muove lentamente!).
Ruolo del nucleo striato:avviene la condensazione del movimento pianificato e in preparazione. Da
qui avverrà la reinformazione dell'area 4, da cui partirà il fascio piramidale.
I nuclei della base si interfaccia con la corteccia, il cervelletto con la periferia: ci sono grossi fasci di
fibre collegate ad esso, che lo informano del movimento che si sta compiendo.
Il cervellletto è impegnato invece nell' "on going" del movimento, riceve info dalla periferia mentre
si sta eseguendo una azione.
Pag.182
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 11
Il circuito dei nuclei della base è costituito da 8 settori, ognuno con una funzione specifica.
Si parla anche di circuito diretto e indiretto, sempre a livello dei gangli della base.
Ansa lenticolare: efferenza dai nuclei, proietta al talamo con il suo fascio ascendente, con il suo
braccio discendente proietta alla reticolare, e questo va a controllare il tono reticolare sui gamma
che è alla base della regolazione posturale. La stessa via sia stabilizza il corpo quindi, sia attiva il
movimento.
Il talamo:pag.184
Nuclei del complesso ventrale: VPL VPM, di natura sensoriale somatica, VA VL VM invece di
riproiezione motoria, sono di convergenza della via cerebello-talamo-corticale (CBL
nell'immagine). Questi ultimi ricevono anche dall'ansa reticolare.
VA è diretto in particolare alle aree premotorie, mentre VL a quelle motorie:in lesione del VA si
verificano problemi di pianificazione del movimento, veceversa in lesioni al VL si verificheranno
problematiche all'esecuzione del movimento in se.
I nuclei intralaminari:
vi appartiene per es il Centrum Medianum, intervengono intervengono nel ritmo sonno veglia,
nella sincronizzazione della corteccia, servono per distinguere tra dolore e nociocezione, (ossia
l'oggettività o la soggettività del dolore). Il dolore verrà poi trasmesso al sistema limbico.
CM proietta anche ai nuclei della base, ecco quindi perchè una forte emozione può causare
ipercinesia.
Circuiti della corteccia:pag.183-194
Sono 4 tipologie, disposti in parallelo, separati tra loro, deputati a controllo e organizzazione di:
1.Funzioni scheletromotorie.
2.Regolazione dell'oculomozione e della foveazione (frontal eye fields)
3.Funzioni associative
4.Il sistema limbico
I sistemi striato-pallidari sono organizzati a clusters di neuroni, come se le 4 vie appena analizzate
siano collegate a gruppi, a clusters appunto.
In dettaglio:
1.Circuito associativo:
origina da corteccia, (prefrontale aree 9-10, premotoria 6, parietale posteriore 7), caudato, pallidus,
substantia nigra, VA e DM.
2.Circuito oculomotore:
controlla il sistema oculomotorio: 1 via diretta ai collicoli superiori, importante per pianificare il
movimento.
3.Circuito limbico:
viene coinvolto il nucleo accumbens, che appartiene al sistema limbico, avrà un ruolo comunque
importante nel movimento, in particolare correlato alle emozioni.
Le vie del SEPOC sono glutamatergiche, e proiettano a putamen e accumbens.
I neuroni detti spinosi, dello striato, (molto grandi, come i motoneuroni alfa e le cell di Purkinje del
cervelletto), sono modulati da sostanza nigra pars compacta, che produce dopamina: l'unione tra
dopamina e glutamato permette una modulazione. Lo striato è anche stimolato dal sepoc.
(Il morbo di Parkinson è causato da una mancata regolazione dei sistemi in esame. Tutti i farmaci
parasimpatico mimetici potrebbero aumentare la rigidità parkinsoniana).
Lo striato, il caudato il putamen proiettano al globus pallidus, distinto in uno interno e uno
esterno:
Il circuito diretto è costituito dal pallido interno (azione, movimento), viceversa quello indiretto è
costituito con il pallido esterno (stop, fine dell'azione): utilizzano GABA, inibitore fondamentale
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 12
per il talamo: il pallido è inibente sul talamo, ma se inibisco una inibizione avrò una attivazione.
Con una sola via si potrà sia attivare il sistema, sia inibirlo.
Pag.192
Nel circuito diretto si avrà: inibizione dell'inibizione: (-)(-) = + del pallido su talamo.
Invece in quello indiretto: GP esterno è inibito-->nucleo subtalamico stimola il globus pallidus
interno e reticolare che vanno ad inibire il talamo (di solito il GP ext inibisce il nucleo subtalamico.)
In caso di decerebrazione, la midbrain extrapiramidal area (MEA) non riceve più le efferenze del
pallido, inibenti la MEA stessa-->scaricamento della MEA nei gamma motoneuroni-->spasticità.
A queste vie si deve aggiungere anche la via iperdiretta dello stop:
corteccia-->stimola il nucleo subtalamico-->stimolazione dell'inibizione al talamo, che sarà
bloccato immediatamente.
Pag.188
Disturbi e problematiche correlate:
Ipercinesie: corea di Hungtinton:causato dal fatto che il pallidus interno non riceve più inibizione
dal nucleo striato-->si avrà una minor inibizione del talamo. Problematica correlata al GABA.
Oppure esiste l'emiballismo.
Bradicinesia: MdP, morbo di Parkinson: dovuto a difetti di dopamina, nonchè alla riduzione
dell'inibizione sul g.pallidus. Il pallido interno inibisce maggiormente il talamo, mentre il pallido
esterno non inibirà più il n.subtalamico, che ecciterà il pallido interno.
Talamo: inibito in modo molto consistente dal pallido interno.
197-202, 204:pagine riassuntive più importanti
Patologie associate:
1.Il morbo di Parkinson:pag.212-216
Patologia che coinvolge la substantia nigra (di Sommering), caratterizzata da tremore a riposo,
difficoltà di fermarsi e ipocinesia. Si distinguono:
1.Sintomi primari: tremori, rigidità, instabilità posturale, bradicinesia
2.Sintomi secondari: crampi, depressione, micrografia, ipomimia, disartira...
Cure:
1.Levodopa: principalmente, ma ha dei lati negativi (necessità di aumento continuo di dose, perdita
di efficacia)
2.DBS: deep brain stimulation:
3.Trapianti: in scimmie si è provato ad autotrapiantare il glomo carotideo, ricco di dopamina. Pare
funzioni con tali animali, non si è tentato nell'uomo.
Oppure, ma non si fa, si potrebbe tentare il trapianto di ghiandole surrenali.
Infine, si è prelevata sostanza nera da embrioni abortiti, e pare ci sia uno sviluppo corretto della
substantia nigra, senza rigetto (attenzione: in realtà dovrebbe avvenire lo stesso il rigetto, nonchè
sviluppo di teratomi...).
Pag.203
Il tremore parkinsoniano e il talamo:pag.250-261 (da vol I)
I neuroni talamici possono "oscillare" come durante il sonno, passano da tonici a oscillanti.
Dipende da una ridotta produzione di dopamina nella substanzia nigra (pars compacta)-->nucleo
striato-->nucleo subtalamico (tramite GABA)-->globo pallido (tramite GLUT). Si ha poi un input
abnorme dai nuclei della base ai nuclei talamici VA, VM e VL-->area 4: tremore a riposo, che è
spiegato quindi dal fatto che le scariche ritimiche si proiettino alla corteccia motoria.
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 13
I nuclei talamici considerati sono di convergenza tra output del pallido e del cervelletto: anche a
seguito di lesione di cervelletto si può avere un tremore, ma in movimento. Viceversa il tremore
parkinsoniano si verifica in caso di riposo, da fermi. Il cervelletto si interfaccia con la periferia, più
il soggetto si muove e più il deficit si manifesta. Mentre nel parkinsoniano ciò è dovuto a
dipendenza dalla corteccia, a una stretta interfaccia con essa.
• Si può raffreddare il nucleo dentato del cervelletto, sperimentalmente-->si avrà un tremore
nel movimento. I neuroni oscilleranno, come durante il sonno.
Cura:
1.nel tremore parkinsoniano si è visto che è possibile interrompere l'oscillazione tramite
talamotomia delle zone coinvolte-->ma si avrà una degenerazione valleriana e pure interneuronica,
intersinaptica, si ingigatisce la lesione così: si avranno danni sensitivi e nella fonazione, nella
parola.
2.Più comodo è l'uso della L-dopa, ma è necessario somministrare di volta in volta dosi sempre
maggiori.
3.DBS: deep brain stimulation:nucleo subtalamico di Lewis: sperimentalmente si è visto che
quando si causa elettrocoagulazione, avviene esaurimento dei neuroni, il nucleo si "spegne" -->fine
del tremore. Si inserisce un apparecchio apposito sottocute, e si collega un filo con elettrodo che
termina nel nucleo subtalamico.
Pallido interno: da uno stimolo GO al movimento, quello esterno invece NO-GO.
Scarica tonica: sotto influsso di Na, mentre scarica oscillante: sotto Ca.
2.01.2010
2.I tremori:pag.223-233
sono i motoneuroni che sono comandati ad oscillare, dal "tremor oscillator": l'oscillazione è
causata da talamo, nuclei della base, cervelletto, corteccia.
In ogni caso saranno coinvolte le vie piramidali:i nuclei talamici passano da una scarica continua a
una oscillante-->proiettano a fascio piramidale, e da qui ai motoneuroni. Ci sono diversi tremori,
che si studieranno in neurologia.
Il tremore parkinsoniano (dei gangli della base):
Nel Parkinson aumenta la ritmicità talamica, infatti i neuroni talamici non sono più ben coordinati
dai sistemi di uscita dei nuclei della base: quando i nuclei VA VL VM non ricevono correttamente
gli stimoli dall'ansa reticolare, trasmetteranno in maniera intermittente.
Ci sono 3 elementi principali in tale circuito alla base dell'oscillazione: nuclei talamici specifici,
corteccia cerebrale, formazione reticolare talamica (che fa oscillare il talamo).
I neuroni talamici possono modificare il loro stato:
1.Attività tonica: interviene il Na.
2.Attività ritmica: intervengono burst di Ca.
Tale attività talamica sarà correlata all'attività dell'organismo:
1.Veglia: Na, attività tonica
2.Anestesia/sonno: burst di Ca, attività ritmica (un barbiturico potrà inibire l'attività del sistema
reticolare e talamico: si verificherà tale stadio di burst).
I nuclei sono però correlati alla corteccia, si verificherà quindi che nel sonno, tali nuclei sono
sincronizzati. Nel sonno la corteccia ha attività intrinseca, cessano molte attività e impulsi diretti
alla corteccia stessa. Eventuali imput (rumori, ansie...) possono interrompere il sonno e generare
scariche continue nel talamo.
Il tremore parkinsoniano dipende dall'acquisizione dei neuroni talamici di una ritmicità intrinseca,
questo perchè diventano privi del controllo dei nuclei della base, a loro volta privi di dopamina,
fondamentale per regolarli.
Un tempo, tramite la talamotomia funzionale era possibile ridurre i tremori (VA VL VIM), ma
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 14
inevitabilmente si aveva anche una ipotonia.
E possibile però in maniera più precisa stimolare maggiormente il talamo, non distruggendo i
neuroni ma facendo consumare loro il neurotrasmettitore. L'elettrodo viene posto sempre a livello
del talamo, e il paziente in caso di tremore poteva attivare il macchinario per fermare tale
movimento involontario.
Il tremore cerebellare: senza il cervelletto, o con sua lesione, il sistema non è anticipatorio, ossia
non c'è una anticipazione del movimento, ma si basa sulle informazioni afferenti che provengono
dal movimento (per es quando sta per cadere un oggetto): si verificherà una oscillazione tra la
posizione reale e quella desiderata.
Sono coinvolti soprattutto le lesioni ai nuclei dentato e interposito:in questo caso quindi si
verificherà che:
esecuzione del movimento-->percezione dell'atto-->inviato stimolo di correzione-->continuo
inseguimento tra correzione e movimento: il tremore si manifesterà in modo crescente più ci si
avvicina a un obiettivo: tale tremore avviene durante il movimento ed è detto quindi volontario,
viceversa nel Parkinson avviene a riposo, è detto non intenzionale.
Operazioni al cervello-Craniotomie:pag.234-243
Asportazione di una porzione di osso ("come uno sportello"), sotto di cui si troveranno le meningi.
Rimozione del tumore avviene per suzione tramite una pompa; prima di rimuovere il tessuto
maligno si stimolano le aree (a cui corrisponderà un movimento nel paziente, secondo le varie
mappe).
Prima dell'intervento si esegue un esame attento della struttura cranica, infatti si opera in base alla
struttura specifica del paziente. Il cranio viene fissato tramite apposite strutture, si esegue la
stereotassi.
Importante è anche stabilire i cosidetti punti di rèpere, in modo tale da localizzare le strutture più
interne: si individueranno i diversi piani, detto Frankfurter, interauricolare (meato acustico ext-
arcata orbitaria inferiore, oppure più comunemente ci si basa sulla linea bicommissurale: linea che
idealmente unisce commissura anteriore e posteriore).
Un altro piano di riferimento è quello cantomeatale, passante per il forame acustico e rima
palpebrale. Serviranno anche le altre coordinate facilmente individuabili.
Verranno eseguite anche radiografie, risonanze magnetiche, computerizzate e studiate,
sovrapposte tra loro: si otterrà una griglia-atlante tipica di ogni paziente.
(CFR fis I che ha spiegato la stessa cosa)
• Cellule staminali nel cervello: totipotenti, possono essere indotte a sostituire i neuroni
malati: sperimentalmente si è visto che avviene una migrazione di cell per riparare il danno
che si è verificato. Si potrebbe utilizzare tali sistemi per es nel Parkinson, trasformando tali
cell in neuroni dopaminergici.
In conclusione:pag.217-221
• Pars reticolare di Sub.Nigra + Globus pallidus int si possono considerare come un unicum
• Sub.nigra pars reticolata pare sia coinvolta soprattutto nella via verso il collicolo superiore
piuttosto che nella via del moto.
• Lesione a via discendente: perdita di inibizione a formazione reticolare-->iperattività
gamma e rigidità.
• Stimolazione di striato, nel circuito indiretto, provoca inibizione dei movimenti spontanei e
posturali = a lesione al pallido esterno-->acinesia
• Lesione caudato e putamen-->ipercinesia.
• Tramite il VA gli stimoli si dirigono a corteccia cerebrale; aree associative e cognitive, sono
sia colpite nei pazienti parkinsoniani, sia causa di demenza.
• Tramite recenti studi si pensa che i nuclei della base non generino il movimento, ma
inibiscano i muscoli antagonisti, questo perchè pare che tali strutture inibiscano le azioni
che andrebbero contro le azioni volontarie.
• Riassunto organizzazione del movimento: piano+preprogramma-->programma, elaborato
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 15
grazie a vie piramidale/cervelletto. La copia efferente resta a livello del cervelletto--
>esecuzione (motoneurone alfa, placca motrice...)-->correzione se necessario, paragone tra
inteso ed eseguito.
• Parametri del movimento: posizione (tramite recettori articolari); forza (recettori tendinei);
velocità (esplorativa, al buio, o balistica, veloce, alla luce); accelerazione (carezza o
schiaffo).
Il sistema limbico; i sistemi della memoria:pag.251-289
Riferimenti anatomici:pag.251
• Ippocampo: fondamentale nell'apprendimento cosciente (studio, cultura...), mentre il
cervelletto in quello incosciente.
• Cervelletto ed ippocampo sono cortecce a 3 strati, c'è una analogia tra loro, sono delle
"macchine", capaci di elaborare e riciclare degli eventi, e poi immagazzinarli.
• I circuiti dell'emozione e quelli della memoria cosciente ed esplicita sono sovrapponibili, gli
eventi che generano emozione si ricordano più facilmente.
• A questo ultimo punto si ricollega il circuito di Papez, appartenente al sistema limbico, che
include anche l'ippocampo. E' costituito da 2 uscite, una cerso il mesencefalo, che permette
di rispondere subito a una emozione, è la via del "go"; un'altra diretta verso la corteccia
(aree 24-25 in particolare e dirette poi alle aree 11 e 47): via del " no go" , della riflessione.
Questo per sottolineare il link tra memoria ed emozione.
Circuito di Papez:
http://www.neuropsicologia.it/img_sezioni/ruggeri/fig01.jpg
Il sistema limbico:
Pag.254
L'ippocampo: suddivisione anatomica: subiculum, presubiculum, parasubiculum... corno di
Ammone...Il giro dentato è un riavvolgimento modificativo di strutture della circonvoluzione
temporale inferiore: da una corteccia a 6 strati si passa a una da 3, la tipica corteccia ippocampale.
Tramite il fornice si porta ai nuclei mammillari, mentre il fascio mammillo-talamico è diretto ai
nuclei talamici-->si va poi verso il giro cingolato e poi di ritorno all'ippocampo.
Tutte le strutture connesse ad ippocampo (ma anche al cervelletto) ricevono pure fibre dallo stesso,
c'è una certa reciprocità quindi-->ciò sarà fondamentale per la ripetizione dell'evento nei circuiti
neurali.
Amigdala: in essa si individuano vie afferenti ed efferenti. Essa è responsabile come detto degli
stimoli emozionali, per es o sottocorticali talamici diretti, di reazioni non coscienti quindi, correlate
alla paura per es. Permette risposte rapide, talvolta anche pericolose, perchè non c'è l'intervento
corticale. Senza di essa non ci sarebbe l'apprendimento, dato il sottolineato nesso tra emozione e
apprendimento. Situazioni di edema cerebrale possono portare a compressione di amigdala.
Riceve ed invia stimoli:
input da esterno, da visione o comunque stimoli esterni-->afferiscono a talamo e corteccia--
>output,
Lo stimolo può o arrivare prima di tutto al talamo, in modo tale che si scateni una risposta
improvvisa, prima della percezione dello stimolo, oppure arrivare in una via "alta", corticale, che
permette la comprensione e l'elaborazione dello stimolo.
Amigdala è connessa anche con ippocampo:nelle persone che hanno subito un evento di violenza,
la memoria di tale evento resta perenne: l'amigdala è attivata nell'evento, e proietta all'ippocampo:
ecco il perchè del ricordo è persistente.
Amigdala quindi viene attivata nel ricordo dall'ippocampo, mentre dall'altro lato, una volta
attivata, stimola nuovamente l'ippocampo, duplice connessione.
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 16
(L'ippocampo si attiva per connessioni reciproche, sono presenti circuiti riverberanti (a un impulso
inviato corrisponde un altro impulso indietro).
Essa è importante anche nella relazione tra madre e figlio: infatti le afferenze olfattive sono
strettamente connesse al sistema limbico e ad amigdala: come sappiamo tali stimoli non hanno un
relay talamico, ma arrivano direttamente al rinencefalo.
Pag.287:pathway dello stress: lo stimolo inviato da amigdala può tornare indietro, è retrogrado,
grazie a NA, rediretto anche a ippocampo.
Pag.287
La paura: pag.262
Sarà caratterizzata da tremore, pallore, aumento della freq cardiaca.
Efferenze di amigdala: verso tutte le strutture implicate nella paura:
Ipotalamo-->attivazione del sistema simpatico
Nucleo motore dorsale del vago-->attivazione del parasimpatico, perdita del tono degli sfinteri,
riflesso di incontinenza a causa della paura, oppure si può avere il fenomeno della fuga dal vago,
arrestando il ritmo cardiaco.
Centro apneustico-->che controlla la frequenza respiratoria viene attivato notevolmente.
Locus coeruleus, nucleo laterale dorsale del tegmento, area tegmentale ventrale-->produzione di
Ach, NA, dopamina.
Nucleo paramentricolare: stimolazione della produzione di ACTH-->produzione a livello surrenale
di ormoni dello stress/ altri-->ritenzione di Na, aumento pressione arteriosa...
(ecco perchè fattori di stress sono pericolosi per la salute).
Lobo dell'insula: si osserva aprendo la scissura di Silvio, è poco studiata perchè difficilmente
raggiungibile, è importante sede di afferenze olfattive e gustative. Connessa comunque al sistema
limbico, correlata a eventi come l'imprinting, studiato molto da Konrad Lorenz (vi prende parte
anche ippocampo).
Siamo noi che creiamo la paura, è una risposta ad una situazione che si intuisce possa essere
pericolosa. Si verificherà: freezing (sistema antico per sfuggire ai predatori, fingendosi morti),
aumento della pressione grazie all'ipotalamo laterale, stress (con produzione di ormoni correlati,
grazie al nucleo paramentricolare, fondamentali per riadattare l'organismo in tale situazione, come
ACTH e adrenalina, NA... ), riflesso di allarme, grazie per es al ponte, al caudato...
Apprendimento e visione:
Visual learning: l'apprendimento avvienetramite la visione del colore, della forma, delle
caratteristiche degli oggetti e del mondo che circonda il bambino.
Le afferenze visive arrivano al lobo occipitale: si verificherà una sorta di "stratificazione
successiva" delle informazioni:forma generica-->limiti, contorni-->colori: prima vediamo in bianco
e nero e subito dopo avviene "la colorazione".
Si parla poi anche della dicotomia della visione: una tipologia di visione è deputata all'azione:
quando si guarda un oggetto, si stabilisce sia la distanza per es dello stesso (oppure le
caratteristiche come dimensione...): tali segnali arrivano ad aree motrici, infatti da qui scaturirà il
movimento.
Un altra visione è fondamentale per la comprensione: si studierà le caratteristiche spaziali,
cromatiche etc...di un oggetto.
Lesioni che colpiscono la prima via causeranno agnosia visiva, ossia un'alterata coscienza spaziale
degli oggetti, mentre quelle che colpiscono la seconda causeranno prosopagnosia, ossia il non
riconoscimento di volti conosciuti.
Il lobo temporale sarà importante per la conoscenza dell'ambiente, e per la memoria: interconnette
infatti, nel processo di memorizzazione, l'amigdala e l'ippocampo. Se leso, si verificheranno perdite
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 17
mnemoniche.
Esiste anche l'apprendimento involontario, che avviene tramite il cervelletto, al quale si affida
l'apprendimento di azioni ripetitive, come camminare, guidare...
• Importanza dell'emozione nel comportamento umano:sensazione/percezione-->emozione
scatenata-->memoria che si sviluppa a seguito di tali eventi-->azione.
• Anosagnosia: negazione di malattia, tipico di casi di demenza avanzata.
• Le prime memorie che si formano sono di tipo motorio.
• Il "cervello tripartito":Paleocervello-Archicervello-Neocervello: i più "antichi", il
neocervello, il più recente. Sembra che non ci sia però, per lo meno a livello funzionale, una
divisione così stretta.
13.01.10
La memoria:pag.263-289
Working memory: ossia la memoria che si spende nel breve termine, per es pochi minuti, sfrutta
conoscenze pregresse. (perdere il filo del discorso è per es un segno di poca working memory).
A tale memoria affluiscono afferenze di amigdala (contenuti emozionali) ma anche di ippocampo
(memorie pregresse).E' diversa dalla cosidetta memoria a breve termine.
E' localizzabile a livello della corteccia frontale/prefrontale.
Saranno implicate diverse strutture: le conoscenze pregresse sono collocate a livello ippocampale,
mentre l'attività emozionale è connessa all'amigdala.
Lettura a voce alta: facilita la memorizzazione, grazie alle afferenze acustiche:
Afferenze acustiche-->paraippocampale-->entorinale-->vie perforanti-->fascia dentata-->Mossy
fibers-->cell CA3-->tramite collaterali di Shefferd torna a CA1 quindi subiculum ed entorinale--
>giro perippocampale-->neocortex. Pag.265
Le informazioni alla fine sono depositate nell'area 38 del lobo temporale. (asporatazione bilaterale
di tale aree causa perdita di tutte le info del passato, si parla di amnesie retrograde). E' comunque
coinvolto anche il lobo frontale:in esso si trovano aree motoria, premotoria (6,8), prefrontale, ossia
quella di più recente evoluzione, di convergenza di elementi razionali (provenienti dal talamo), e di
elementi emozionali (da sistema limbico).
• Errori di sommazione della memoria:errori riguardanti la memoria associativa: se si
presentano 2 stimoli vicini nel tempo,senza che la memoria si consolidi, si verificheranno
sovrapposizione di nomi, concetti, immagini. Ciò non succederà se c'è stato il tempo per
consolidare una memoria.
• Ippocampo: deputato alla memoria esplicita, cosciente, dichiatativa, non procedurale,
mentre l'amigdala a memoria non dichiarativa, procedurale.
• Fonological loop: inteso come loop dello stimolo visivo-uditivo; permette di mutare, far
evolvere un discorso, permette che lo stesso sia coerente, con una struttura ordinata.
Tipi di memoria:
Pag.272
• Memoria cosciente: quella che fa parte delle memorie stabilite secondo apprendimento, di
tipo cosciente. (anche detta declarativa o esplicita).
• Memoria non cosciente:molte memorie sono di tipo non cosciente, in particolare più si
continua ad eseguire una operazione e più tali atti faranno parte di tale memorie.
• Memoria esplicita:cosciente e non procedurale, suddivisibile in memoria episodica,
riguardante gli eventi della vita, oppure memoria dei fatti, semantica, correlata alla
capacità di associazione tra parole, idee, concetti. A questo si ricollega la capacità di parlare:
da bambini si apprende sentendo i suoni, poi dai 6 anni si comprende anche il concetto
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 18
insito nella parola, e infine da adulti l'apprendimento è strettamente cosciente.
Ad essa è connesso il cosidetto apprendimento non associativo, in cui lavora una memoria
esplicita che associa eventi: questo avviene nell'imprinting, che è una sorta di
apprendimento istintivo, non c'è un programma cosciente. Avviene anche nell'adattamento,
in cui c'è una sorta di imprinting negativo (noia, in cui uno stimolo ripetitivo fa diminuire
l'attenzione e la memorizzazione).
• Memoria non dichiarativa, non cosciente e procedurale , classificabile in diversi ambiti:
competenze motorie e abitudini, priming (è il fenomeno dei messaggi subliminali, non è
ancora ben compreso:interviene nella pubblicità, quando vengono proiettati per brevissimo
tempo dei fotogrammi, che non sono visti consciamente ma inconsciamente, e che
comunque fanno aumentare l'interesse per quell'oggetto).
• Memoria e linguaggio: la lingua madre viene appresa in modo non cosciente,
"passivamente". Tali memorie vengono depositate nell'emisfero sinistro, anche se il
bambino apprende più di una lingua:si parla in questo caso di bilinguiscmo compatto.
Tali lingue "si accumulano" a strati, quindi a seconda dell'entità di un danno cerebrale (per
es ictus) si possono perdere una o più lingue. Le lingue apprese da adulti invece sono
depositate nell'emisfero destro (bilinguismo acquisito).
La localizzazione della memoria:pag.274
A livello di lobi frontale e temporali in particolare, ma in ogni caso ci sarà un continuo crearsi e
disfarsi di connessioni tra i neuroni, una variazione continua dei circuiti neuronali e dei legami
neurochimici.
Sembra che nella memoria siano implicati anche i nuclei della base, ma non si sa bene come e in
che misura (probabilmente il nucleo caudato è implicato in memoria esplicita).
Cervelletto: di sicuro è implicato, proietta in tutta l'area 6.
Amigdala: uno stimolo che si porta a tale struttura può trasformarsi in un output, diretto a :
corteccia sensoriale: si avrà una memoria a breve termine; ippocampo: memoria a lungo termine;
corteccia prefrontale: working memory.
Pathway del cosa e del dove:
Pag.279
In tali immagini sono raffigurate le cosidette pathway del cosa e del dove, nei confronti della
visione e della working memory.
E' coinvolto un sistema ventrale, del "cosa", in cui è implicata la corteccia temporale, che riconosce
l'oggetto, nonchè la corteccia visiva che permette di percepire lo stimolo visivo.
Inoltre è coinvolto un sistema dorsale, in cui è implicata la corteccia parietale per la localizzazione
dell'oggetto, nonchè la corteccia prefrontale.
Tutto in ogni caso converge alla corteccia prefrontale (aree 11,12,46,47): bisogna dettagliare però
che: le porzioni mediali permettono una memoria a breve termine, mentre quelle laterali e inferiori
danno una memoria a lungo termine. Tale corteccia prefrontale è connessa con il basal forebrain,
che agisce tramite Ach (è costituito da benderella di Brocà, dai nuclei del setto...).
Ippocampo:presenta elevata concentrazione di recettori NMDA, per le memorie esplicite e
permanenti, che saranno depositate in area 38.
Si avrà prima di tutto attivazione dei canali AMPA non NMDA-->depolarizzazione che diffonde
elettrotonicamente ai canali NMDA-->apertura dei canali e ingresso di Ca2+-->si scatena pathway
con calmoduline e si attiva adenilato ciclasi, stimolata anche da dopamina (facilitante la
memorizzazione), e Ach (stesso effetto, grazie al nucelo di Meynert). (Serotonina invece inibirà la
memorizzazione). Avviene poi attivazione delle MAP kinasi, e successivamente i TF CREB 1 e 2,
proteine leganti cAMP, fosforilati. Attiveranno sintesi proteica coinvolta in memorizzazione.
Le amnesie:pag.275
1.Amnesia anterograda: causata da lesione ippocampale, non c'è sviluppo di nuove memorie,
restano quelle passate.
2.Amnesia retrograda: implica lesioni ad aree 38 per es, vengono perse le memorie passate ma
sono create quelle recenti.
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 19
3.Sindrome di Korsakoff: amnesia retrograda progressiva, notata soprattutto negli alcolisti, si
perdono prima i fatti più vicini e poi quelli lontani, sempre più lontani...E' possibile che sia dovuta
anche a lesione a livello dei corpi mammillari: sembra che partecipino a una progressiva perdita di
memoria.
• Il fenomeno del condizionamento semplice classico: si distingue uno stimolo assoluto (per
es mostrare a un cane una ciotola di cibo), e uno neutro (suono di campanella), non
associato direttamente al primo stimolo: sarà l'esperienza ad associare insieme i 2 stimoli
differenti, tanto che il cane dopo un po' sarà pronto a mangiare (secrezione di saliva) anche
solo al suono della campanella.
14.01.10
Il cervelletto:pag.290-348
Riferimenti anatomici-introduzione:pag.298-300
Fondamentale per il controllo del movimento, oltre ad essere coinvolto nella memorizzazione
implicita.
Ma non si è definito con chiarezza dal pdv dei compiti: è caratterizzato infatti da una attività tonica,
ad alta frequenza, la cui funzione potrebbe essere o tonigena o necessaria per garantire una minima
attività motrice.
In ogni caso una lesione al cervelletto darà un movimento poco coordinato o sregolato; compare
dismetria, ma in ogni caso il movimento permane.
Si sviluppa inizialmente come struttura vestibolare: tali nuclei evolvono dai pesci (quindi i
primigèni nell'evoluzione), fino a noi.
• Fondamentale è la capacità di arrestare il movimento sul target: il cervelletto deve regolare
quindi sia inizio sia fine.
• Il cervelletto è inevitabilmente implicato nello stato di veglia (movimento) ma anche di
riposo (non movimento).
Nell'animale:
Se si asporta cervelletto nel gatto, si ha animale decerebellato: il cervelletto nel gatto invece è
inibente su strutture vestibolari-->si avrà una rigidità di tipo alfa.
Sempre in un animale decerebellato si verificherà una iperestensione a livello nucale.
Nell'uomo:
Il cervelletto ha funzione facilitante (effetto Rossi) su vie reticolo spinali: se asporto, i neuroni
vengono disfacilitati,e quindi non avrò più funzionamento del sistema gamma-->comparsa di
ipotonia.
Il cervelletto regola si il movimento ,ma ha anche un ruolo tonigeno, è fondamentale questa sua
azione, (tramite vie cerebello-reticolo-talamiche). E' importante come sappiamo nel controllare gli
atti motori automatici e abitudinari. Da' tono in oltre alle vie piramidali-->fluidità, tramite via
cerebello-talamo-corticale.La lesione di tale struttura porta alla disgregazione dell'anello alfa-
gamma. Il paziente cerebellare non avrà più una fluidità nelle azioni.
• Lesione nuclei della base: scatena ipertono gamma, perchè hanno effetto su vie reticolo
spinali maggiormente.
• Nell'uomo il cervelletto è costituito per il 90% da neocerveletto, per il 10% da
paleocervelletto.
Pag.51-52:
• Il cervelletto è disposto non in serie sui sistemi motori (se spezzo un anello della catena, la
catena non va più), è disposto in parallelo. I peduncoli lo uniscono al bulbo e al ponte.
• Una lesione del cervelletto non darà paralisi, ma un movimento atassico e asinergico.
• Le cell di Purkinje sono biplanari, sono estese notevolmente, ma in 2 dimensioni quasi, in
tal modo ce ne stanno di più, sono impilate.
• Cuneo cerebellar tract (CCT): originato dal nucleo cuneato, (nucleo di Burdach: ha anche
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 20
proiezione a cervelletto).
Classificazione:
Diviso a seconda di diversi criteri:
criterio filogenetico:
archicerebello (lobulo flocculonodulare, che riceve anche afferenze dirette dal vestibolo,
dall'utricolo, dal sacculo...),paleocerebello (10%) (funzioni motorie di locomozione,
movimento...),neocervelletto (90%). Si stratifica, è una progressiva stratificazione.
Divisione longitudinale:zona vermiana, paravermiana e laterale: tale divisione è più consona alle
vie di entrata.
mentre quella trasversale prende in considerazione di più i sistemi di ingresso idem (lobus
anterior posterior e flocculonodulare).
Divisione funzionale: vestibolocerebellum, comunicante con i nuclei vestibolari, spinocerebellum,
correlato al midollo spinale, e pontocerebellum, in relazione con corteccia con talamo da
intermediario (VA e VL ricevono anche con una proiezione detta dentato talamo-cervelletto).
Struttura fine:
Nuclei cerebellari:
Dentato, emboliforme, globoso,fastigio, in più quello di Dayters, considerato il 5 nucleo cerebellare
(benchè appartenente ai nulei vestibolari), riceve afferenze dal cervelletto.
Ad attività tonica, aiutati dalle collaterali delle fibre muscoidi e rampicanti: saranno responsabili di
una sorta di "livello minimo" del motore, caratterizzata da una scarica ad alta frequenza, tonica
appunto.
Corteccia fatta da 3 strati (dei granuli, del purkinje e molecolare), 5 tipi di cellule: granuli, golgi,
purkinje a canestro e stellate, e 2 tipi di fibre: rampicanti e muscoidi: c'è una unita, uniformità
strutturale.
Cellule:
1.Di Purkinje: enormi, le più grandi di tutto l'encefalo, con conformazione bidimensionale ("come
una mano"), possono disporsi una dietro l'altra. L'unica via di uscita sono gli assoni delle cell di
purkinje-->finiscono nei nuclei intracerebellari, tranne quello di Dayters: dentato (parvicellulare,
ventrale-->VL-->area 4; magnicellulare, dorsale-->VA-->aree premotorie), interposito (proiezione
al nucleo rosso e al talamo), fastigio (anteriore e posteriore-->nuclei intralaminari talamici):
tramite esso si attiva il centro vascolare (in preparazione al movimento).
2,3.Cellule a canestro/stellate: avvolgono le cell di P.
4.Golgi: attivate dalle fibre parallele, esercitano inibizione sulle cell dei granuli e quindi le fibre
muscoidi, di conseguenza.
Analogia con i nuclei della base: su 5 tipi di neuroni, 4 sono inibitori, gabaergici, si avrà come nei
nuclei della base l'inibizione dell'inibizione. Inibizione: tramite GABA. Solo le cell dei granuli sono
eccitatorie.
Folia: microcirconvoluzioni, che aumentano di molto la superfice cerebellare (area totale: 0,75 mq,
quasi come il cervello). E' lamellato quasi. Il folium ha una proiezione trasversale, si espandono in
senso mediolaterale, mentre le cell del purkinje sono perpendicolari a queste: si incrociano di 90°.
Fibre e vie:pag.320-322
(studiale)
Proiezioni pontocerebellari: 20 milioni di fibre entrano nel cervelletto. I nuclei pontini ricevono
dalle vie fronto, parieto,temporopontine, tutti i lobi degli emisferi proiettano qui:
Frontale: perchè si programma il movimento
Parietale:perchè movimento modifica percezioni sensitive
Temporale: perchè il movimento può essere una risposta a stimoli uditivi
Occipitale: perchè il movimento piuò essere una risposta a visione
In questo modo il movimento diventa quindi qualcosa di più che una semplice attività di moto.
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 21
Più si espandono gli emisferi, più si espandono anche le vie, c'è una perfetta corrispondenza e
proporzionalità.
Il ponte sarà fondamentale per: inviare a cervelletto fasci da tutta la corteccia e per ricevere da
cervelletto, inviando a corteccia, tramite il talamo a proiezione diffusa.
Come anticipato, il cervelletto invia e riceve fibre da una struttura, c'è una doppia via (concetto di
reciprocità delle vie del cervelletto).
Fibre di ingresso:
1.Fibre muscoidi: si originano tutte dalle vie di ingresso (ponto, reticolo cerebellari...). Tali fibre si
articolano nei granuli e creano le fibre parallele, dette così perchè sono parallele all'asse maggiore
del folium. Sono disposte su un piano trasversale del cervelletto, come i folia, perpendicolari alle
cell del Purkinje.
Fibre rampicanti: correlate a una descrizione trasversale, originano solo dal nucleo olivare
inferiore. Tutte le altre vie sono di tipo muscoide. Tali fibre si chiamano così perchè si arborizzano
con l'albero dendritico delle cell del purkinje. Il piano è perpendicolare all'asse del folium.
In tal modo si vede come le 2 vie di input si intersecano tra di loro di 90 gradi:
muscoidi: parallele ad asse, fondamentali per il circuito del go.
rampicanti: perpendicolari ad asse , fondamentali per il circuito del no go.
Tale disposizione ricorda una scacchiera (si parla della cosidetta scacchiera neuornale): tale
incrocio sarà importante per il controllo reciproco.
Quando gli stimoli muscoide e rampicante si incontrano, si verifica la massima attività cerebellare:
nella zona intorno a tale incrocio, i due stimoli si elidono vicendevolmente-->è proprio in tale
modo probabilmente che c'è il controllo dei nuclei sottostanti (si parla di strisce di attivazione).
Fibre in uscita:
Unica via d'uscita sono le cell di P. : ecco perchè tutti gli altri input sono inibitori.
Verso corteccia:
Si ha attivazione prima delle cell di P, e poi delle cell a canestro (asinaptica per P, monosinaptica
per cell a canestro): ne consegue prima, tramite cell di P. una inibizione dei neuroni nucleari, che di
norma, grazie all'attività tonica cerebellare, facilitano il movimento. Se sono inibiti, come in questo
caso, si avrà una disfacilitazione. Successivamente intervengono le cell a canestro, inibitorie le cell
di P. , aumenta l'attività tonica del nucleo cerebellare, causando una facilitazione talamica.
Circuiti implicati:
1.Circuito del go: corteccia premotoria-->regioni pontine-->fibre muscoidi-->eccitazione granuli--
>inibizione di Purkinje tramite cellule a canestro e stellate-->fine inibizione nuclei.
(più correttamente si è visto che i granuli attivano prima per un istante le cell di P. E poi attivano
cell stellate e basket, inibenti le cell di P.).
2.Circuito del no go: cell di golgi bloccano fibre muscoidi-->fibre rampicanti stimolano le cell di
p.-->inibizione diretta dei nuclei.
• Si deduce che le cell di Golgi regolano l'input delle fibre muscoidi, tramite un apposito
sistema a feedback (cioè retroattivo).
• Inibizione: GABA. Cell a canestro: più potenti perchè agiscono sul soma, meno potenti
sono le stellate, perchè agiscono sui dendriti.
• Altri sistemi inibenti il movimento (correlati al no go quindi) : Locus coeruleus e
adrenalina: controllo del calcio intracellulare, elemento fondamentale per la
memorizzazione. Rafe magno: produce serotonina, importante induttore del sonno-->si
avrà una riduzione dell'attività tonica del cervelletto.
Tali neuromediatori entrano nel cervelletto.
Fasci spinocerebellari: pag.53-65
(DScT, dorsal spinocerebellar tract e ventral ST): il primo porta info da quando l'individuo
appoggia il piede a terra, mentre il VScT segnala fenomeni flessori, la fase legata al programma
centrale. Le afferenze profonde al cervelletto sono sempre ipsilaterali, il fascio dorsale diretto: è già
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 22
ipsilaterale, quello ventrale proviene dall'altra metà del midollo, si incrocia e dvienta ipsilaterale.Il
cervelletto discrimina l'azione grazie a tali vie, e agli impulsi che mandano. Sono sempre
ipsilaterali. 1 è sotto i collicoli inferiori, 1 vicino ai capi midollari.
• Gatto inserito in una struttura con campo a barre:si fa ruotare il gatto mentre vede il campo
a strisce. L'altro vede passivamente le strisce, e da qui si è capito che il bambino costruisce il
suo mondo cosciente nell'esplorare attivamente il mondo. Se si studiano le aree visive del
gatto attivo, avrà aree organizzate, mentre l'altro non avrà tale organizzazione.
Altre funzioni e danni al cervelletto:
1.Cervelletto come comparatore:
fondamentale per controllare e raffinare i movimenti, compara gli input con gli output:
decisione di muoversi-->parte stimolo da corteccia-->copia efferente e nuclei pontini: c'è ancora
una idea del movimento, non sono ancora in movimento-->arriva al cervelletto, che riceve
"l'inteso"-->stimolo intanto arriva al motoneurone, si attivano i fasci spinocerebellari-->tramite i
fusi neuromuscolari e l'organo di Golgi arriva al cervelletto lo stimolo "eseguito": il cervelletto
eseguirà la comparazione.
L'eseguito (via spinocerebellare) deve essere comparato con l'inteso (via pontocerebellare): alla fine
si avrà un'azione che combacia sia nell'eseguito che nell'inteso. Entrambe le afferenze al cervelletto
venogno veicolate dal peduncolo cerebellare medio.
Tale azione comparativa è importante anche nell'apprendimento: movimenti corretti saranno
dovuti alla sovrapposizione perfetta tra inteso ed eseguito.
Se invece il movimento non è avvenuto in modo corretto, le vie dentato-talamo-corticali riferiscono
questo errore alla corteccia (cervelletto come circuito riverberante).
2.Cervelletto e tempo: Il movimento è ovviamente legato al tempo, il cervelletto segue una certa
coordinazione temporale, altrimenti i movimenti sarebbero imprecisi ("il cervelletto è un
temporizzatore");
anche il linguaggio ha una compontente in cui è importante la scansione temporale: sembra che il
cervelletto sia quindi anche depositario di tale scansione temporale.
Un paziente cerebellare perde anche tale capacità, tale cognizione temporale, nonchè quella di
parlare correttamente, avrà delle disartrie (frase ideata e sintatticamente corretta ma senza
prosodia, monotona). In più non è capace di trasferire il sostantivo nel corrispondente fatto di
moto (forchetta-->mangiare, scarpa-->camminare...). Viene distrurbata anche l'idea, l'ideazione
del movimento.
Riassunto funzioni:
1.Controllo dei riflessi vestibolari
2.Controllo dei circuiti spinali
3.Programmazione, inizio, controllo, comparazione del movimento
Cervelletto e alcolismo: in caso di etilismo cronico la corteccia dal pdv biochimico è diversa, viene
più colpito il verme, il paraverme:c'è una diversa capacità da parte delle cell di P. Di utilizzare la
vitamina B6 (tutte le vitamine B sono neuroprotettive): gran parte della corteccia perde le sue
funzioni, colpendo soprattutto le 2 zone indicate sopra. Alcolismo distrugge anche nucleo
fastigiale. Si avrà anche una differente sensibilità alla vitamina B, ci sono quindi alla base delle
differenze biochimiche. Uno dei segni più evidenti dell'alcolismo è l'atassia, nonchè alterazione
dello stato mentale.
• Lesione del fascio piramidale darà paralisi flaccida.
• Rispetto delle leggi di Murray (il raggio al cubo della sezione madre è uguale al raggio al
cubo della somma delle sezioni figlie) e Fitt (più fine è il movimento, più lento è il
movimento: questo non sempre, vedi nell'atletica leggera o nei musicisti).
• Confronti tra animali: uomo:15 mil di cell su 60 Kg di media di peso. Gatto: 3 Kg e 1 mil e
500 mila fibre. Topo: 300 g, 340 mila fibre. Facendo dei rapporti il ratto è l'animale più
capace dei 3 di muoversi al buio, con rapidità...Anche gli uccelli hanno grandi abilità di
movimento.
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 23
• Il cervelletto dei musicisti professionisti sembra più sviluppato del 5%, questo per motivi di
trasporto di ioni e richiamo di acqua: non c'è un aumento delle cellule, ma è un indice di
maggior attività. C'è anche un effetto legato agli ormoni sessuali (si verifica tale fenomeno
soprattutto nel maschio).
19.01
Ablazione:pag.323-324
Ablazioni sono compiute su primati, nell'uomo il corrispettivo saranno le lesioni cerebellari
(benchè le differenze funzionali siano notevoli, basti pensare al neocerevello che occupa solo il 50%
nei primati e il 90% nell'uomo, e ciò favorirà il manifestarsi delle lesioni).
1.Ipotonia:perdita della facilitazione del cervelletto sulle vie cerebello reticolo spinali: è una
diminuzione dell'anello alfa gamma.
2.Astenia:ossia debolezza, causata dalla mancata azione del fascio piramidale (che determina la
forza del movimento in associazione al cervelletto-->se manca azione tonica cerebellare su
corteccia-->si ha perdita di tono).
3.Dismetria: alterazione delle sinergie tra gruppi di muscoli agonisti e antagonisti:movimento
impreciso e alterato, sempre di più, man mano che ci si avvicina al target (per es invece che toccarsi
la punta del naso ci si tocca la guancia).
4.Adiodococinesia: il paziente non riesce a spostare la mano in posizione supina o prona.
5.Atassia: andatura barcollante, "da ubriaco". Allargamento della base di impianto per
muoversi/andamento indeciso per danni vestibolari.
6.Lesioni ristrette:per es a causa di tumori: per es nistagmo (rapidi movimenti oculari, a riposo,
senza stimolo), cisti vermiane, (cavità riempite di liquido che danno atassie).
7.Disartrie: diverse dalle afasie. Nei disartirici la frase è corretta ma priva di prosodia.
8.Tremori:dovuti a differenze tra la posizione desiderata e quella effettiva, reale-->danni
cerebellari non consentono il controllo a feedback del movimento.
Tali patologie si manifestano differentemente, in base alla sede colpita:
1.Archicerebellum: viene alterato l'equilibrio
2.Paleocerebellum: atassia, o disturbi di deambulazione.
3.Neocerebellum: ipotonia, asinergia, astenia, disartria...
Esperimenti di stimolazione del cervelletto:pag.325-329
se si stimola corteccia, si inibiscono i nuclei sottostanti (e non avviene invece loro stimolazione, che
era dovuta ad eccessive correnti erogate durante gli esperimenti).
1.Dentato: interviene nella programmazione del movimento, fondamentale nella via dentato-
takamo-corticale: esperimento con scimmie: prima di un loro movimento si attiva il nucleo
dentato, e sono attivi anche durante il movimento stesso. Lesione a tale nucleo causano ritardo
della partenza del movimento, una disfacilitazione dei neuroni corticali, nonchè un allungamento
del tempo di scarica dei muscoli agonisti.
2.Interposito: scarica durante il movimento.
3.Fastigio: si attiva alla fine del movimento, associato a fenomeni correttivi. I suoi neuroni
codificano alcuni per la forza, altri per la velocità del movimento (2 grandezze inv prop.).
I sistemi sensoriali:pag.349
Fondamentali per il corretto funzionamento dell'organismo. Si possono considerare dei trasduttori
analogico-digitalici, ossia trasformano gli stimoli dell'ambiente esterno in altri stimoli
comprensibili dall'encefalo. Facendo ciò esplicano la loro funzione di trasduttori.
Non si possono sempre discriminare con facilità, infatti per es il dolore dovuto a pressione è
causato da una iperstimolazione dei recettori tattili, oppure la percezione tra caldo e freddo
dipende dalla frequenza di scarica (caldo: aumenta, freddo: diminuisce)...
Dal PDV anatomico saranno coinvolti:
1.Fascio piramidale: importante per il controllo degli input, si aggiunge ai centri superiori in questa
funzione.
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 24
2.Talamo: una sorta di filtro, fondamentale per il relay delle afferenze (tranne per olfatto), verso la
corteccia.
Da ricordare la differenza tra sensazione e percezione: la prima è più rapida, è il ricevere uno
stimolo, la seconda è rendersi conto dello stesso, è più lenta.
La sensibilità somestesica:pag.353
1.Sistemi anterolaterali spinotalamici o extralemniscali: sistema termodolorifico tattile e
superficiale; è protopatico, ossia ci avverte di uno stimolo doloroso.
Il sistema di movimento che è associato ad esso è quello extrapiramidale.
2.Sistemi lemniscali: compaiono in un secondo momento dal PDV evolutivo; è epicritico, ossia
permette di comprendere con precisione e finezza uno stimolo (permette anche di comprendere a d
occhi chiusi un punto di contatto (stereognosia)). In oltre si individua la sensibilità vibratoria o
pallestesia, oppure la cinestesia. Il sistema di movimento associato è quello piramidale.
Nel sistema lemniscale sono coinvolti in particolare 3 neuroni: 1.ganglio spinale, 2. neurone del
nucleo di Goll e Burdack, dello stesso lato, e 3.neurone talamico.
Tra il nucleo di G e B e il talamo si troverà l'incrocio lemniscale. Inoltre, la cosidetta laminazione di
tale nucleo è precisa e somatotopica.
I dermatomeri:pag.354-355
sono delle zone del corpo a cui è associato a livello sottostante un preciso nervo spinale-->si potrà
rilevare facilemente una lesione o una anestesia a livello di un preciso (e univoco) neuromero.
Il concetto di campo recettoriale:
è la capacità di distinguere 2 stimoli come separati dal PDV dello spazio: la sensibilità del corpo
verso stimoli esterni è diversa in base alla zona considerata (polpastrelli, lingua: elevata sensibilità;
gamba, braccio, schiena: sensibilità molto minore).Questo dipenderà dal numero di recettori
presenti in una determinata zona: essi poi invieranno gli stimoli alle struture citate
precedentemente e quindi alla corteccia, in cui sarà presente una precisa mappa dell'homunculus
sensitivus (più recettori-->maggior superfice corticale riservata a tale zona).
Le aree somatosensitive:pag.358-362
1.Primaria:a livello della circonvoluzione parietale ascendente, suddivisibile in porzioni:
area 3A riceve informazioni dai fusi neuromuscolari, la 3B dai recettori cutanei a lento
adattamento, l'area 1 dai recettori a rapido adattamento mentre l'area 2 dai recettori cutanei
profondi.
Ci sarà una sorta di raffinamento progressivo degli stimoli, a partire da area 3a fino alla 2, in modo
tale che l'info venga integrata, determinando così natura, localizzazione, movimento dello stimolo:
tale informazioni convergono poi all'area 5, in cui ci sarà una coordinazione.
2.Somatica secondaria: all'interno della scissura di Rolando, a livello sempre del lobo parietale. E'
un'area superassociativa, in cui afferiscono stimoli cutanei, acustici, visivi.
Il corpo calloso avrà un ruolo in tali sistemi: esso, con i suoi 800 mil di fibre (soprattutto
inibitorie), è capace di integrare tra loro zone simili dal PDV della topografia. In base alle zone del
corpo considerate, ci saranno connessioni callosali differenti (da notare come in caso di sezione del
corpo calloso non si verificano stimoli coordinati).
La surrounding inhibition:pag.363-366
Pressione applicata a un punto della cute stimola anche zone limitrofe, come una sorta di rumore
di fondo, che non permette, se non inibito, di percepire correttamente lo stimolo: sarà la zona
centrale nei nuclei di Goll e Burdach che inibisce le zone laterali, i contorni dello stimolo.
Sono ridotti i rumori di fondo, il segnale è amplificato così, perchè si aumenta la differenza tra
rumore di fondo e stimolo.
20.01.10
Sensibilità termodolorifica:pag. 367
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 25
Sensibilità protopatica, i neuroni deputati a tale sensibilità sono terminazioni libere, hanno campi
periferici ampi (tranne che per il dolore puntorio). Ci sono dei fasci anterolaterali (lemnisco
spinale) dove il 2 neurone della via è collocato nel corno posteriore del midollo (sostanza gelatinosa
di Rolando).
Il dolore:368
Coinvolge diversi centri talamici (nuclei intralaminari): centrum medianum, centralis medialis,
parafascicularis, che proiettano ad aree del sistema libico, all'insula e lobo prefrontale.
Si distingua quindi la nocicezione e dolore: la prima è oggettiva, la seconda soggettiva, è più di una
semplice sensazione, è inevitabilmente compresa anche una componente emozionale.
E' fondamentale per la sopravvivenza, serve a farci capire che c'è qualcosa che non va, è una spia
d'allarme, un prodromo a eventi più gravi. E' quindi, purtroppo, utile.
Distinto in somatico e viscerale: questo ultimo è relativo agli organi, ai visceri appunto: si parla di
dolore riferito (riferito alla cute sovrastante, al dermatomero): questo perchè i visceri non hanno
una rappresentazione a livello dell'homunculus sensitivus: è possibile che tale dolore derivi da
afferenze provenienti sia dal viscere ma anche dalla cute-->proiezione a stesso neurone, che si
porta a livello encefalico, e poi riproietterà a cute (per il fatto che non c'è homunculus appunto).
Oppure si parla di dolore infiammatorio, in quanto nella sede di infiammazione avviene la
produzione di serotonina, istamina, prostaglandine (che causano tumor rubor dolor calor, functio
laesa ): tale dolore deriverà dalla sensibilità chimica delle terminazioni libere (non capsulate).
Infine il dolore ischemico è causato dalla mancaza di O2-->produzione di acido lattico e altre
sostanze che abbassano il pH-->stimolazione delle fibre nervose periferiche, sempre libere e
sensibili a tali sostanze.
Il dolore somatico è diviso in superficiale (caratterizzato da una doppia natura: puntiforme,
precocemente) e + diffuso, in un secondo momento, dovuto questo ultimo all'attivazione delle fibre
del IV tipo di Lloyd, + lente, e profondo, implicato per es nei crampi (che si manifestano a livello
dei muscoli scheletrici in assenza di O2-->produzione di acido lattico e poca ATP-->i filamenti
actomiosinici resteranno legati tra loro; in più gli ioni H+ causano una depolarizzazione e una
contrazione crampiforme). Può riguardare anche articolazioni, oppure essere presente in caso di
emicrania (aumento del volume del contenuto della scatola cranica-->tensione a livello delle
meningi e dolore).
Le vie di conduzione del dolore:
le fibre provenienti dai recettori periferici sono capaci di raggiungere un solo neurone, c'è una certa
convergenza delle afferenze-->il risultato di questo fatto è che la sensibilità dolorifica è sia diffusa,
ma anche altamente imprecisa. Si dice che il campo periferico del neurone dolorifico è molto
ampio. Tutto questo perchè non è tanto importante comprendere la zona precisa che è stata
danneggiata, quanto è fondamentale invece allontanarsi dalla noxa, generando anche una risposta
di tipo fight or fly.
Ci saranno 4 vie dolorifiche afferenti ai centri superiori:
1.Neospinotalamica: implicata nella percezione del dolore, con maggior precisione; associata a VPL
e VPM.
2.Paleospinotalamica: coinvolta nel dolore in modo più generico, proietterà a: nuclei
intralaminari-->zone del sistema limbico, corteccia frontale, lobo insula.
3.Spinoreticolare: vi si distingueranno vie reticolospinali discendenti, vie spinoreticolari ascendenti
(correlate agli eventi di allarme).
4.Spinomesencefalica: coinvolta nella manifestazione a parole del dolore (urla, imprecazioni...) e
nel controllo centrifugo del dolore (gate control). (Esso è anche implicato in una via discendente:
eccita il nucleo di Onuf durante l'atto sessuale, ipedendo fuoriuscita di urina).
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 26
Controllo del dolore:
Oppiacei endogeni e molecole analgesiche:
1.sostanze importanti per il controllo del dolore; dipendono anche da fattori emotivi. Derivano
dalla POMC (proopiomelanocortina) e sono comprese endorfine, encefaline, oppiacei endogeni,
prodotte a livello del sistema limbico, grigio periacqueduttale...
Tale sostanza è capace di far uscire ione K+ dalle cell nociocettrici e quindi iperpolarizzarle (=
inibirle).
2.Alcool: permette la liberazione di endorfine, ha un effetto simile alla morfina. Fu usato come uno
dei primi analgesici.
3.Morfina: analgesico agente su vie dolorifiche. Si lega ai cosidetti recettori κ,μ,δ,γ-->attivazione
dei canali al K+ e iperpolarizzazione del neurone.
Il gate control-la teoria del cancello:pag.375-378
Il dolore non passa sempre a livello dell'encefalo, può essere "fermato" da tale sistema:dopo un
trauma, e alla percezione del dolore, ci si tocca e massaggia subito la zona colpita, provando
sollievo: questo perchè a livello del corno posteriore del midollo è possibile una gate, c'è un
"cancello", una "porta", che blocca il dolore, tramite l'intervento della sostanza gelatinosa di
Rolando.
In tale fenomeno le fibre propriocettive possono inibire presinapticamente quelle sottili, vicarianti
il dolore:
fibre dolorifiche-->arrivano alla sostanza gelatinosa, in cui si trova il 2 neurone della via, che, se
attivo, si porta all'encefalo tramite la colonna anterolaterale. A questo punto può essere importante
la presenza della fibra sensitiva di diametro maggiore presente a livello del sistema lemniscale, può
inviare delle collaterali che raggiungeranno la sinapsi tra il 1 e il 2 neurone della via analizzata-->la
trasmissione verrà inibita.
Tale teoria è una estensione della teoria classica di Hidd:nella sifilide terziaria (che portava a danni
a cordoni posteriori) i pazienti avevano dei fortissimi dolori. Ciò è dovuto alla cosidetta tabe
dorsale: è come una situazione neurodegenerativa,lede i cordoni posteriori, c'è una lesione del
sistema lemniscale.
Pag.376
Controllo discendente:pag.385-389
centrale, che si aggiunge al gate: basato fondamentalmente sul PAG e sulle encefaline:
PAG:attivato, insieme al rafe, dallo stimolo dolorifico-->attivazione di una vie discendente che può
coinvolgere il locus coeruleus (serotoninergico, Nadrenergico)-->stimolazione di neuroni
encefalinergici.
Encefaline:controllano l'afferenza dolorosa iperpolarizzando le cellule portatrici dello stimolo:i
recettori agli oppiacei (recettori κ,μ,δ,γ) saranno capaci di:
a.aprire i canali al K+
b.chiudere i canali al Ca
Così la cellula sarà iperpolarizzata e meno eccitabile.
• Esiste anche un controllo legato alla volontà dell'individuo (controllo cognitivo).
• Anche il rafe può controllare il dolore, oltre ad essere fondamentale per regolare il sonno
(ecco perchè c'è un legame tra anestesia e sonno indotto).
• Pratiche spesso molto discusse come l'agopuntura avrebbero una base tramite questa teoria
del cancello.
La sensibilità termica:pag.390-394
Recettori capaci di percepire la temperatura. Si pensa che, dal PDV fisiologico, tali sensazioni siano
dovute a una differente scarica (freddo: calo della frequenza, caldo: aumento), mentre dal PDV
anatomico si ritiene che ci siano recettori differenti (di Krause per il caldo e di Golgi per il freddo).
Ad ogni modo la percezione del freddo è più facile, diffusa, perchè il freddo è più pericoloso per
l'uomo.
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 27
Tali strutture sono ad adattamento lento, infatti se fossero rapidi non permetterebbero di percepire
un danno, un pericolo.
Probabilmente il caldo è associato all'apertura di canali al Na, mentre il freddo è associato a
chiusura di canali al K.
Inoltre la scarica sarà differente, varierà la tempistica (il K si accumula lentamente, il Na entra
molto velocemente).
I sensi
L'udito:pag.395-439
Senso che permette all'uomo di percepire le afferenze sonore provenienti dall'ambiente, tra cui le
parole per esempio, dei fonemi che hanno un significato specifico.
Nell'uomo il range di frequenze udite va da 20 Hz a 20 kHz. Le frequenze più piccole ("i bassi"), si
propagano con maggiore facilità. (Frequenza: 1/T, è il numero di oscillazioni compiute nel tempo).
Il suono viene prodotto a livello delle corde vocali, grazie alla loro vibrazione e tensione: si
verificherà una modulazione nella frequenza e nell'altezza.
La conformazione dell'arcata laringo-faringo-buccale è stata fondamentale per lo sviluppo della
fonazione nell'uomo.
Gli Hz raggiunti nell'uomo sono:200-2500 nel maschio, 200-3500 nella donna: questo grazie alla
diversa conformazione delle strutture ossee nei due sessi.
Leggi fisiche e concetti che regolano i suoni:
1.Intensità o ampiezza: è la potenza del suono. La somma di suoni con stessa frequenza non fa
aumentare la frequenza ma l'intensità.
2.Energia totale di un suono:dipende da: mezzo di propagazione (più è solido, e più si trasmette il
suono); dalla frequenza e dall'intensità.
Con K=costante del mezzo analizzato, P=pressione, omega=freq.
3.Potenza del suono:misurata in decibel (dB): tale valore è uguale a dieci per il Log del rapporto
I/I0 (I=intensità):decibell = 10log(I/I0).
Da notare che i valori sono espressi su scala logaritmica, quindi ci saranno delle notevoli differenze
in termini di dB, anche se può sembrare il contrario...
(0dB: strappo di un foglio; 100,110 dB: danni a strutture dell'orecchio medio).
4.Leggi:la capacità di percepire i suoni è regolata soprattutto da 2 leggi:
a.Legge di Weber:"uno stimolo deve essere aumentato di una frazione costante del suo valore
affinchè la diversità sia percepibile".
b.Legge di Fechner:"una sensazione provocata da uno stimolo aumenta di una quantità costante
quando lo stimolo viene aumentato di un fattore costante.
Riferimenti anatomici:
Diverse strutture collaborano alla percezione e localizzazione sonora:
1.Orecchio esterno: il padiglione auricolare: ha il compito di amplificare il suono (senza di esso il
suono calerebbe di 20 dB)
2.Naso/viso: influiscono sulla percezione, localizzazione dei suoni, sono degli ostacoli anche per il
suono, e contribuiscono comunque alla stereofonia-->il ritardo che causano sarà alla base della
localizzazione sonora.
21.01.10
Orecchio medio:
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 28
3.Timpano: strutturato "ad imbuto", ha una superficie maggiore rispetto alla finestra ovale. I 3
ossicini, staffa, incudine e martello, collaborano all'amplificazione del segnale uditivo.
Orecchio interno:
Strutture dell'orecchio interno
Pag.401
collocato a livello dell'osso temporale: vi si trovano soprattutto 2 strutture fondamentali per la
percezione sonora (coclea) e per il sistema vestibolare (utricolo, sacculo, canali semicircolari).
Ai 3 ossicini si associano anche dei muscoli, con una peculiare funzione amplificante.
La finestra ovale: trasmette le vibrazioni all'endolinfa, si colloca in contatto con la staffa. E' 20-30
volte più piccola del timpano, e riceve un suono amplificato già di 20 dB.
La finestra rotonda:fondamentale per far "scaricare" l'energia pressoria che si è accumulata
precedentemente: è capace di espandersi se la pressione è eccessiva.
Le 2 finestre collaborano nella trasmissione dell'onda.
La tuba di Eustachio: serve per il controllo pressorio a livello di entrambe le facce del timpano
(ecco perchè in caso di "orecchio tappato", per es in aereo, l'atto dello sbadiglio può ripristinare il
livelli di pressione).
Pag.405
• Otiti: possono essere pericolose, soprattutto se croniche, infatti se non curata può evolvere
in meningite, data la stretta vicinanza di tali strutture con zone del lobo temporale e
appunto le meningi.
• Fenomeno di Tullio: ossia la possibile interverenza tra il sistema vestibolare e quello
cocleare-->modificazioni delle afferenze acustiche possono portare a modifiche delle
afferenze vestibolari.
Tale sistema ha il compito di percepire le vibrazioni sonore, che poi verranno convertite in stimolo
elettrico diretto alla corteccia.
Le vibrazioni possono essere distinte in:
1.Vibrazione aerea: a livello delle prime vie acustiche, padiglione in particolare
2.Vibrazione meccanica: trasmessa grazie agli ossicini
3.Vibrazione idrodinamica: a livello dell'orecchio interno
e infine la trasduzione (e trasmissione con solita modalità elettrochimica).
La coclea (pag.406-409 immagini) è caratterizzata dalla tonotopia, ossia la percezione di certe
frequenze sarà tipica di una sua specifica area piuttosto che un altra.
Al suo interno si trovano: l'organo del Corti e diverse strutture cellulari, tra cui per es le cellule
ciliate, all'interno dell'organo del Corti:sono dotate di stereociglia.
Presenta poi la membrana basale, che si allarga seguendo lo sviluppo della coclea: ciò è alla base
della tonotopia cocleare:alla stimolazione della coclea si genera un'onda che è caratterizzata da un
cosidetto ventre e un apice, quest'ultima è importante nella percezione-->è capace di inibire le
frequenze attorno ad essa (con un fenomeno simile alla surrounding inhibition).
Un suono con elevata frequenza ed energia si esaurisce subito con l'impatto, viceversa uno con
frequenza bassa si protrae all'interno della struttura: quindi le prime porzioni della membrana
trasmettono le frequenze alte, e viceversa le ultime, di dimensioni maggiori.
Struttura fine: caratterizzata da 3 scale:
1.Vestibolare: il suo liquido, la perilinfa (ricca di Na e con poco K), riceve le vibrazioni generate
dalla staffa.
2.Cocleare: in cui si trova l'organo del Corti. Presenta un liquido, l'endolinfa, con alti livelli di K e
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 29
meno di Na, prodotto dalla stria vascolare, un tessuto molto vascolarizzato, caratterizzato da
numerose pompe Na/K con azione invertita.
3.Timpanica:comunica con la finestra rotonda, sempre per far diminuire la pressione.
Pag.407
Meccanismo di trasmissione:
Il suono, con le sue vibrazioni, causa le vibrazioni della perilinfa -->vibrazione della membrana
basale-->si verifica poi il fenomeno della trasduzione meccanoelettrica, grazie alle ciglia delle
cellule capellute, in contatto con la membrana tectoria (che si trova sopra le cell stesse, che sono
anche immerse nell'endolinfa)--> lo stimolo meccanico eccita le cell capellute, fa aprire i canali al
K-->ingresso di K (220 meq/L) in tali cell (generazione del potenziale endococleare o
microfonico)-->depolarizzazione e produzione di stimolo elettrico, che viaggia tramite il nervo
acustico. Il potenziale di riposo sarà ripristinato con uscita di Na da cell.
Le cell capellute hanno una funzione connessa alla loro disposizione: quelle interne sarebbero
coinvolte nella percezione del suono, quelle esterne invece pare servano per raffinarelo stimolo e/o
controllare la percezione del suono-->ci saranno delle connessioni con delle vie efferenti al SNC
(fascio olivococleare di Rasmussen) che moduleranno l'organo del Corti-->si elimineranno , come
in una sorta di filtro, le frequenze non desiderate.
Le vie acustiche:410-433
Caratterizzate da numerose stazioni, nonchè da vie ipsilaterali e controlaterali (+ spesso, sono il
triplo delle ipsilaterali ): in ogni caso saranno attivate entrambi gli emisferi corticali.
Stazioni/nuclei:410-413
1.Ganglio del Corti
2.Nuclei cocleari
3.Corpo trapezoide
4.Complesso olivare superiore
5.Nuclei del lemnisco laterale
6.Collicolo inf.re
7.Genicolato med.le
Da qui gli stimoli si portano alla corteccia.
Nuclei cocleari: importanti nella surrounding inhibition. Anche a questo livello sarà rilevabile una
organizzazione tonotopica (presente anche nel genicolato med.le).
Collicolo inferiore: da esso si dipartono le vie collicolospinali, che garantiscono il riflesso acustico-
motorio (la risposta istintiva di fronte ad una afferenza uditiva).
Nuclei olivari: proiettano a cervelletto e ai collicoli (ipsilaterali e controlaterali ).
• Tale complessità di vie ha dei nessi con l'evoluzione: infatti, la fine capacità di distinguere
suoni, singole parole...potrebbe derivare dalla necessità che avrebbero avuto i nostri avi nel
difendersi e percepire anche il minimo suono.
Le zone corticali:pag.414
Le aree corticali deputate alla ricezione degli stimoli uditivi sono collocate a livello del lobo
temporale (41-42 in particolare, giro di Heshl, a livello delle circonvoluzioni superiori), deputate
alla percezione ed elaborazione dell'input.
Sono principalmente 3, con la tipica disposizione tonotipica di tale via (A1: ant.te: freq.
Elevate;post.te: freq. Basse; A2: disposizione invertita; A3 ).
In tali lobi si può verificare una notevole asimmetria:infatti, il lobo sn (in cui sono presenti anche i
centri del linguaggio) è più esteso del dx: tali zone di sn avviene un ottimale riconoscimento della
frequenza (più precisamente in A1 che in A2). Inoltre in tale zona le cellule sono molto grandi ma
poche, numericamente.
Nelle zone di dx invece avverrà la comprensione della prosodia dei suoni; non ci sarà una
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 30
tonotopia; le cell sono piccole e ben più numerose che a sn-->ciò permette probabilmente di
analizzare i suoni dal pdv spaziale.
Controllo della percezione sonora:415-423
centrifugo, discendente, mediato dal fascio olivococleare di Rasmussen (riguardante il nucleo
olivare superiore). Anche la corteccia è implicata nel controllo (tramite vie ipsi/controlat.):
il fascio si porta a livello delle cellule esterne dell'organo del Corti, grazie a fibre di grande diametro
(controlaterali) e di diametro minore (omolaterali)-->si avrà una modulazione periferica della
coclea, ossia avverrà una inibizione dei "rumori di fondo" e una esaltazione del suono principale.
Inoltre anche i muscoli degli ossicini (mm stapedio e tensore del timpano) permettono una
amplificazione del segnale: questo perchè, per es nella fonazione, tramite i nuclei del trigemino e
del faciale è possibile modificare la tensione timpanica-->aumentarne la capacità trasmissiva--> in
tal modo l'ascolto della nostra stessa voce (parola, canto...) è favorito rispetto ad altri stimoli uditivi
. (in ogni caso noi sentiamo la nostra voce anche grazie alla risonanza delle ossa craniche).
L'audiogramma-psicologia e udito:pag.424426
Test eseguito per determinare la capacità uditiva di un individuo: si potranno analizzare le risposte
dell'apparato a diverse intensità e frequenze:
max sensibilità: 1600 Hz (frequenza udibile però solo con intensità che raggiunge i 60 dB)
il limite inferiore saranno i 16 Hz (udibili solo a 120 dB).
Il campo per la parola si collocherà ovviamente entro tale delimitazione (rientra infatti in 250-
3500 Hz): l'udito umano sarà tarato e basato su essa.
Pag.424
Teorie dell'udito:
1.Ipotesi di Volley:sperimentale, dimostra che con dei suoni puri (non naturali), le fibre nervose
(presenti in numero di 30-50 mila) si comportino in maniera differente:
a.Stimolo minore o uguale a 300 Hz:le fibre scaricano contemporaneamente
b.Stimolo superiore a 300 Hz, per es 600: le fibre si dividono in 2 gruppi, e scaricano in modo
alternato.
c.Con stimoli ancora superiori la scarica diviene ciclica-->questo porta ad un aumento della
frequenza di scarica che è correlata alla frequenza del suono percepito.
Entra in gioco poi la tonotopia-->l'impulso verrà poi trasferito in base all'intensità con frequenze
diverse.
Gli impulsi trasmettono informazioni quindi relative alla frequenza e all'intensità.
Udito e linguaggio:
• Sordomutismo:secondario alla sordità.
• Un bambino con problematiche uditive a 7-8 mesi non riuscirà a parlare, diventa muto,
questo per il nesso tra udito e linguaggio.
• Si ricorda l'importanza della disposizione anatomica del strutture laringe/faringe e vie
aeree superiori, fondamentale per produrre in modo corretto i suoni che caratterizzano il
linguaggio.
• Come accennato, la posizione del feto nell'utero è importante per la prevalenza destra:
infatti il sistema vestibolare e uditivo sono maggiormente stimolati a dx, nell'utero-->le
proiezioni si porteranno maggiormente all'emisfero sn: in più, l'orecchio destro elimina le
afferenze dirette all'emisfero dx (che riceve da orecchio sn): si parla di ascolto dicotico.
• Tramite diversi esperimenti si è visto che l'orecchio destro è spesso dominante sul sinistro,
per es: si stimolano entrambe le orecchie con delle sequenze (di lettere o numeri)
differenti-->spesso il soggetto analizzato avrà percepito lo stimolo di dx; oppure con la
tecnica del tapping, ossia si fa premere un pulsante per es, con la mano dx-->si stimola
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 31
anche qui entrambe le orecchie, e spesso, lo stimolo di sn non viene percepito: sembra che
ci sia una interferenza tra il movimento e l'udito quindi.
• Aree attivate nella percezione delle parole: aree acustiche 41 e 42; associative 11 e 47;
frontali 8 e 9.
• Aree coinvolte nella fonazione: attivazione aree 39-40 per il pensiero-->area di Vernicke--
>area di Brocà. In ogni caso sarà necessaria una selezione delle parole, una combinazione
corretta delle stesse. (area di Wernike è vicina all'area acustica).
• Turbe del linguaggio: turbe della selezione: sono posteriori, coinvolgono l'emisfero
temporale-->parole non sensate, neologismi. Turbe dell'organizzazione della frase: causate
da disturbi alle aree anteriori.
Un grande ringraziamento va a, in ordine alfabetico:
Giuseppe Marini
Federico Nappi
Giordano Perin
per la loro cortesia e per i loro appunti, con cui ho controllato e integrato le mie
note.
Spero non ci siano gravi errori-ce ne saranno di sicuro (non ho potuto controllare tutto con
precisione), quindi non fidatevi troppo di quello che ho scritto e leggete con una buona
dose di spirito critico :)
Buono studio!
Federico Pippo
Fisiologia II-Vol I-Pippo Federico-A.A. 2009-2010 32
Potrebbero piacerti anche
- Neuroanatomia - Riassunto Del CattaneoDocumento47 pagineNeuroanatomia - Riassunto Del CattaneoAlessandro Fazio50% (2)
- Scaricato Da Internet - Esame Obiettivo in OrtopediaDocumento135 pagineScaricato Da Internet - Esame Obiettivo in OrtopediaOriano Fallace100% (1)
- GnatologiaDocumento20 pagineGnatologiawilliamNessuna valutazione finora
- Diagnostica Per ImmaginiDocumento51 pagineDiagnostica Per ImmaginiFedericoPppzPippoNessuna valutazione finora
- Anatomia 2Documento58 pagineAnatomia 2GioNessuna valutazione finora
- Cervelletto, Nuclei Della Base, Controllo Della Postura, Sistema VentricolareDocumento19 pagineCervelletto, Nuclei Della Base, Controllo Della Postura, Sistema Ventricolareapi-3704348100% (2)
- Il Tuina: La più antica arte manipolativa dei tessuti biologici per il raggiungimento dello stato di buona salute psico-fisicaDa EverandIl Tuina: La più antica arte manipolativa dei tessuti biologici per il raggiungimento dello stato di buona salute psico-fisicaNessuna valutazione finora
- Terminologia ragionata della medicina cinese classica e moderna | Unità 5: Fisiologia di sangue e fluidiDa EverandTerminologia ragionata della medicina cinese classica e moderna | Unità 5: Fisiologia di sangue e fluidiNessuna valutazione finora
- La spalla dello Sportivo: Anatomia e fisiologia della spalla, le disfunzioni da stress atletico (gesto “overhead”) e il loro trattamento riabilitativoDa EverandLa spalla dello Sportivo: Anatomia e fisiologia della spalla, le disfunzioni da stress atletico (gesto “overhead”) e il loro trattamento riabilitativoValutazione: 5 su 5 stelle5/5 (2)
- Algoritmi diagnostico-terapeutici in osteopatiaDa EverandAlgoritmi diagnostico-terapeutici in osteopatiaNessuna valutazione finora
- I Ritmi in Osteopatia: I ritmi all'interno del corpo e nella relazione con l'ambienteDa EverandI Ritmi in Osteopatia: I ritmi all'interno del corpo e nella relazione con l'ambienteValutazione: 5 su 5 stelle5/5 (1)
- Mal di Schiena: Terapia manuale Semeiotica, Diagnosi e tecniche di normalizzazioneDa EverandMal di Schiena: Terapia manuale Semeiotica, Diagnosi e tecniche di normalizzazioneNessuna valutazione finora
- Le correnti a radiofrequenza applicate alla terapia del doloreDa EverandLe correnti a radiofrequenza applicate alla terapia del doloreNessuna valutazione finora
- PROFESSIONE PERSONAL TRAINER - Elementi per una formazione integraleDa EverandPROFESSIONE PERSONAL TRAINER - Elementi per una formazione integraleValutazione: 5 su 5 stelle5/5 (1)
- Il Nervo Vago: La guida completa per capire, stimolare ed attivare il potere curativo ed antistress del nervo vago. Riduci drasticamente ansia, palpitazioni, depressione e molte altre malattieDa EverandIl Nervo Vago: La guida completa per capire, stimolare ed attivare il potere curativo ed antistress del nervo vago. Riduci drasticamente ansia, palpitazioni, depressione e molte altre malattieValutazione: 5 su 5 stelle5/5 (1)
- NeurologiaDocumento226 pagineNeurologiaΑντώνιος Χατζηγεωργίου50% (2)
- NeurologiaDocumento50 pagineNeurologiaSalvatore FerraroNessuna valutazione finora
- Vie SomatosensorialiDocumento4 pagineVie Somatosensorialidimilda100% (1)
- SNP Plesso Cervicale e BrachialeDocumento7 pagineSNP Plesso Cervicale e BrachialeMichelangelo Scanu100% (1)
- Dispensa ReumatologiaDocumento17 pagineDispensa ReumatologiaAlessandro FazioNessuna valutazione finora
- Nervi Cranici FinaleDocumento30 pagineNervi Cranici FinalefedericoNessuna valutazione finora
- Radiologia 1 - 4 AnnoDocumento176 pagineRadiologia 1 - 4 Annobasturk81Nessuna valutazione finora
- 1 - Sensibilità SomestesicaDocumento23 pagine1 - Sensibilità Somestesicaapi-3704348100% (3)
- Fascia CervicaleDocumento14 pagineFascia CervicaleFrancesco Valsega100% (1)
- Muscolo e LocomozioneDocumento21 pagineMuscolo e Locomozioneapi-3704348100% (1)
- Generalità ArticolazioniDocumento27 pagineGeneralità ArticolazioniMatteoDiNassoNessuna valutazione finora
- 5 CranioDocumento6 pagine5 CranioSalvo Danilo LombardoNessuna valutazione finora
- Neurologia GayDocumento136 pagineNeurologia Gaybasturk81Nessuna valutazione finora
- Dispensa NeuroanatomiaDocumento142 pagineDispensa NeuroanatomiaPierpaolo LuzioNessuna valutazione finora
- Malformazione Di Chiari - Tonsille CerebellariDocumento50 pagineMalformazione Di Chiari - Tonsille CerebellarisololuiNessuna valutazione finora
- La Colonna Cervicale, Conoscerla Meglio - Riabilitazione - Atrosi - FisioterapiaDocumento8 pagineLa Colonna Cervicale, Conoscerla Meglio - Riabilitazione - Atrosi - FisioterapiaVico Massagista e Quiropraxia - Massagem, Massoterapia e Ventosa Terapia - São José SC.Nessuna valutazione finora
- Cervicali Sbobbinature-4Documento15 pagineCervicali Sbobbinature-4Sharon TaylorNessuna valutazione finora
- Sistema Gastrointestinale Fisiologia DelDocumento160 pagineSistema Gastrointestinale Fisiologia DelMiriam GalliNessuna valutazione finora
- Vie Afferenti Recettori Tattili, Termici, NocicettoriDocumento12 pagineVie Afferenti Recettori Tattili, Termici, Nocicettoriapi-3704348100% (3)
- Corso RXDocumento140 pagineCorso RXRoberto FranzeseNessuna valutazione finora
- Sistema NervosoDocumento5 pagineSistema NervosoGiuliaNessuna valutazione finora
- Massimi Di Inspirazione, Gas Respiratori e Loro Trasporto, Velocità Di Passaggio Del Sangue Nel Capillare PolmonareDocumento21 pagineMassimi Di Inspirazione, Gas Respiratori e Loro Trasporto, Velocità Di Passaggio Del Sangue Nel Capillare Polmonareapi-3704348100% (1)
- GinecologiaDocumento131 pagineGinecologiaFrancesco SonnatiNessuna valutazione finora
- Funzione MotoriaDocumento19 pagineFunzione MotoriaEdoardo BerteroNessuna valutazione finora
- 36-Anatomia II-16.03.16-Anatomia Strutturistica e MicroscopicaDocumento6 pagine36-Anatomia II-16.03.16-Anatomia Strutturistica e MicroscopicaSandroNessuna valutazione finora
- PolineuropatieDocumento9 paginePolineuropatieapi-3739045Nessuna valutazione finora
- Neuroplasticità e NeuroriabilitazioneDocumento17 pagineNeuroplasticità e NeuroriabilitazionessaNessuna valutazione finora
- Sindrome Dello Stretto Toracico SuperioreDocumento5 pagineSindrome Dello Stretto Toracico SuperioremaxNessuna valutazione finora
- Anatomia e Sindrome Del Piriforme e SciaticaDocumento7 pagineAnatomia e Sindrome Del Piriforme e SciaticaDemetrioIɔɔnʌlɐsNessuna valutazione finora
- Anatomia SNPDocumento30 pagineAnatomia SNPantonio nennaNessuna valutazione finora
- Neuro Chi Rur GiaDocumento29 pagineNeuro Chi Rur GiaGabiţa DorinaNessuna valutazione finora
- Traumatologia e AnatomiaDocumento77 pagineTraumatologia e AnatomiaNicola CreccaNessuna valutazione finora
- Epiteli SensorialiDocumento5 pagineEpiteli Sensorialiapi-3704348100% (1)
- Antonio Rillo Tesi Diploma Osteopatia 2018 PDFDocumento46 pagineAntonio Rillo Tesi Diploma Osteopatia 2018 PDFStudio miofitNessuna valutazione finora
- 5 - Midollo e Riflessi SpinaliDocumento22 pagine5 - Midollo e Riflessi Spinaliapi-3704348100% (2)
- Sinapsi e NeurotrasmettitoriDocumento45 pagineSinapsi e NeurotrasmettitoriNicola Laurora0% (1)
- F 11-02-22 LombalgiaDocumento15 pagineF 11-02-22 LombalgiaAnna FiccoNessuna valutazione finora
- 30 - Anatomia II - 26-04-2017 - RDocumento11 pagine30 - Anatomia II - 26-04-2017 - RSandroNessuna valutazione finora
- Fisiologia Respiratoria (Meccanica, Scambio e Trasporto Dei Gasi, Controllo Nervoso) PDFDocumento13 pagineFisiologia Respiratoria (Meccanica, Scambio e Trasporto Dei Gasi, Controllo Nervoso) PDFBobbyGustafson100% (1)
- RIPASSO Anatomia Umana I Parte PDFDocumento78 pagineRIPASSO Anatomia Umana I Parte PDFDa NyNessuna valutazione finora
- Scienze - Il Sistema NervosoDocumento27 pagineScienze - Il Sistema NervosoRiccardo PengoNessuna valutazione finora
- Sistema NervosoDocumento36 pagineSistema NervosoalinabortaNessuna valutazione finora
- 18 - Anatomia II (Microscopica) - 13.03.2017 - RDocumento8 pagine18 - Anatomia II (Microscopica) - 13.03.2017 - RSandroNessuna valutazione finora
- Stimolo e Risposta Muscolare, RiflessoDocumento16 pagineStimolo e Risposta Muscolare, Riflessoapi-3704348100% (2)
- 2 Sna NeurofisiologiaDocumento34 pagine2 Sna NeurofisiologiaTomyasuNessuna valutazione finora
- Chirurgia EndocrinologicaDocumento24 pagineChirurgia EndocrinologicaFedericoPppzPippoNessuna valutazione finora
- Coltivazione LuppoliDocumento71 pagineColtivazione LuppoliFedericoPppzPippoNessuna valutazione finora
- Anatomia RadiologicaDocumento23 pagineAnatomia RadiologicaFedericoPppzPippoNessuna valutazione finora
- Chirurgia GastroenterologicaDocumento43 pagineChirurgia GastroenterologicaFedericoPppzPippoNessuna valutazione finora
- Medicina NucleareDocumento84 pagineMedicina NucleareFedericoPppzPippo100% (1)
- Aspetti Di Anatomia PatologicaDocumento59 pagineAspetti Di Anatomia PatologicaFedericoPppzPippoNessuna valutazione finora
- ENDOCRINOLOGIADocumento33 pagineENDOCRINOLOGIAFedericoPppzPippoNessuna valutazione finora
- Farmacologia SpecialeDocumento38 pagineFarmacologia SpecialeFedericoPppzPippoNessuna valutazione finora
- PNEUMOLOGIADocumento43 paginePNEUMOLOGIAFedericoPppzPippoNessuna valutazione finora
- Corso Di ElettrocardiografiaDocumento18 pagineCorso Di ElettrocardiografiaFedericoPppzPippo100% (2)
- Fisiopatologia Clinica PT.1Documento22 pagineFisiopatologia Clinica PT.1FedericoPppzPippoNessuna valutazione finora
- ANGIOLOGIADocumento25 pagineANGIOLOGIAFedericoPppzPippoNessuna valutazione finora
- Ematologia IiDocumento39 pagineEmatologia IiFedericoPppzPippoNessuna valutazione finora
- Fisiopatologia Clinica PT.2Documento13 pagineFisiopatologia Clinica PT.2FedericoPppzPippoNessuna valutazione finora
- Malattie InfettiveDocumento43 pagineMalattie InfettiveFedericoPppzPippo100% (1)
- REUMATOLOGIADocumento19 pagineREUMATOLOGIAFedericoPppzPippo0% (1)
- NEFROLOGIADocumento37 pagineNEFROLOGIAFedericoPppzPippoNessuna valutazione finora
- CARDIOLOGIADocumento39 pagineCARDIOLOGIAFedericoPppzPippoNessuna valutazione finora
- GASTROENTEROLOGIADocumento33 pagineGASTROENTEROLOGIAFedericoPppzPippoNessuna valutazione finora
- Fisiopatologia Chirurgica-Chirurgia ToracicaDocumento16 pagineFisiopatologia Chirurgica-Chirurgia ToracicaFedericoPppzPippoNessuna valutazione finora
- Immunologia ClinicaDocumento38 pagineImmunologia ClinicaFedericoPppzPippo100% (1)
- Fisiopatologia Chirurgica PT.1Documento35 pagineFisiopatologia Chirurgica PT.1FedericoPppzPippoNessuna valutazione finora
- DERMATOLOGIADocumento42 pagineDERMATOLOGIAFedericoPppzPippo100% (1)
- Semeiotica Medicina Di LaboratorioDocumento34 pagineSemeiotica Medicina Di LaboratorioFedericoPppzPippo100% (1)
- EMATOLOGIADocumento16 pagineEMATOLOGIAFedericoPppzPippoNessuna valutazione finora
- Chirurgia PlasticaDocumento28 pagineChirurgia PlasticaFedericoPppzPippo100% (1)
- Semeiotica ClinicaDocumento20 pagineSemeiotica ClinicaFedericoPppzPippoNessuna valutazione finora
- Semeiotica Chirurgica-1Documento30 pagineSemeiotica Chirurgica-1FedericoPppzPippoNessuna valutazione finora
- Semeiotica Chirurgica-2Documento37 pagineSemeiotica Chirurgica-2FedericoPppzPippoNessuna valutazione finora
- Domande Da Pinel Gennaio 2012Documento13 pagineDomande Da Pinel Gennaio 2012Anna MaranoNessuna valutazione finora
- LIBRO La Malattia Di ParkinsonDocumento78 pagineLIBRO La Malattia Di ParkinsonAnton SamarxhiuNessuna valutazione finora
- NeurologiaDocumento241 pagineNeurologiaAndrea Rosi100% (4)
- Sistema MotorioDocumento12 pagineSistema MotorioSebastiano67% (3)
- Neur002 1Documento18 pagineNeur002 1gaiaNessuna valutazione finora
- Nuclei Del TelencefaloDocumento1 paginaNuclei Del TelencefaloRolando LagangiaNessuna valutazione finora
- 2.fisiologia II - Neurofisiologia - Parte IIDocumento32 pagine2.fisiologia II - Neurofisiologia - Parte IIFedericoPppzPippoNessuna valutazione finora
- Neurotrasmissione CatecolaminergicaDocumento16 pagineNeurotrasmissione CatecolaminergicaAndrea Grazi100% (1)
- Arrighi Superstar - Fondamenti Terzo Modulo Sistemato DefinitivoDocumento44 pagineArrighi Superstar - Fondamenti Terzo Modulo Sistemato Definitivoluigi cremascoliNessuna valutazione finora
- Neurologia - RiassuntoDocumento38 pagineNeurologia - Riassuntoapi-3863584100% (3)
- Regolazione EmotivaDocumento5 pagineRegolazione EmotivaFabioNessuna valutazione finora
- Scimmie, Ominidi, UomoDocumento46 pagineScimmie, Ominidi, UomoNeuroscienze.netNessuna valutazione finora
- Patologia Generale-Giordano PerinDocumento377 paginePatologia Generale-Giordano PerinGiordano Perin100% (1)
- Domandeleggio CORRETTEDocumento9 pagineDomandeleggio CORRETTEfrancescaNessuna valutazione finora