
Timo
Esta página ou seção foi marcada para revisão devido a incoerências ou dados de confiabilidade duvidosa. |
Este artigo ou secção necessita de expansão. |
| Timo | |
|---|---|
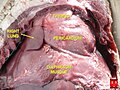
 Localização anatômica do Timo | |
| Detalhes | |
| Sistema | Sistema linfático, parte do Sistema imune |
| Drenagem linfática | Linfonodo traqueobrônquico, Linfonodos paraesternais |
| Precursor | Arco faríngeo |
| Identificadores | |
| Latim | thymus |
| MeSH | D013950 |
| Dorlands/Elsevier | https://meshb.nlm.nih.gov/record/ui?ui=D013950 |
O timo[1] é uma glândula linfóide primária especializada do sistema imunológico. Dentro do timo, os linfócitos T amadurecem. Os linfócitos T são muito importantes para o sistema imunológico adaptativo, onde o corpo se adapta especificamente para combater invasores externos. O timo é composto por dois lobos idênticos e está localizado no mediastino anterior superior, na frente do coração e atrás do esterno. Cada lobo do timo pode ser dividido em uma medula central e um córtex periférico que é cercado por uma cápsula externa.
O córtex e a medula desempenham papéis diferentes no desenvolvimento dos linfócitos T. As células no timo podem ser divididas em células estromais do timo e células de origem hematopoiética (derivadas de células-tronco hematopoiéticas residentes na medula óssea). Os linfócitos T em desenvolvimento são denominadas timócitos e são de origem hematopoiética. As células estromais incluem células epiteliais do córtex e da medula tímica e células dendríticas. O timo fornece um ambiente para o desenvolvimento dos linfócitos T a partir de células precursoras. As células do timo fornecem o desenvolvimento dos linfócitos T que são funcionais e auto-tolerantes. Portanto, um dos papéis mais importantes do timo é a indução da tolerância central.
O timo é maior e mais ativo nos períodos neonatal e pré-adolescente. No início da adolescência, o timo começa a diminuir em tamanho e atividade, e o tecido do timo é gradualmente substituído pelo tecido adiposo (gordura). No entanto, a linfopoiese residual T continua ao longo da vida adulta.
Estrutura
[editar | editar código-fonte]Nas crianças, o timo é cinza-rosado, macio e lobulado em suas superfícies.[2] No nascimento, ele tem cerca de 4 a 6 cm de comprimento, de 2,5 a 5 cm de largura e cerca de 1 cm de espessura.[3] Ele aumenta de tamanho até a puberdade, onde pode chegar cerca de 40 a 50 g,[4][5] o qual diminui de tamanho em um processo conhecido como involução.[5]
O timo é composto por dois lobos que se encontram na linha média superior e se estendem abaixo da tireoide no pescoço até a cartilagem da quarta costela.[2] Os lobos são cobertos por uma cápsula.[4] O timo fica abaixo do esterno, repousa sobre o pericárdio e é separado do arco aórtico e dos grandes vasos por uma camada de fáscia. A veia braquiocefálica esquerda pode até estar ligada ao timo.[2] No pescoço, fica na frente e nas laterais da traqueia, atrás dos músculos esterno-hióideo e esternotireóideo.[2]
Histologia
[editar | editar código-fonte]O córtex é composto principalmente de timócitos e células epiteliais.[4] Os timócitos, células T imaturas, são suportados por uma rede de células reticulares epiteliais finamente ramificadas, que é contínua com uma rede semelhante a da medula. Essa rede forma uma adventícia para os vasos sanguíneos, que entram no córtex via septos próximos à junção com a medula.[2] Outras células também estão presentes no timo, incluindo macrófagos, células dendríticas e uma pequena quantidade de linfócitos B, neutrófilos e eosinófilos.[4]
Na medula, a rede de células epiteliais é mais grossa do que no córtex, e as células linfóides são relativamente menores em número.[2] Corpos concêntricos semelhantes a ninhos chamados corpúsculos de Hassall (também chamados corpúsculos tímicos) são formados por agregações das células epiteliais medulares.[4] São espirais concêntricas e em camadas de células epiteliais que aumentam em número ao longo da vida.[2] São os restos dos tubos epiteliais, que crescem a partir das terceiras bolsas faríngeas do embrião para formar o timo.[6]
-
 Micrografia mostrando um lóbulo do timo. O córtex (área roxa mais profunda) envolve uma medula menos densa e mais leve.
Micrografia mostrando um lóbulo do timo. O córtex (área roxa mais profunda) envolve uma medula menos densa e mais leve. -
 Micrografia mostrando o corpúsculo de Hassall, encontrado na medula do timo.
Micrografia mostrando o corpúsculo de Hassall, encontrado na medula do timo.


Suprimento de sangue e nervos
[editar | editar código-fonte]As artérias que suprem o timo são ramos das artérias torácica interna e inferior da tireóide, com ramos da artéria tireoidiana superior algumas vezes vistos.[3] Os ramos atingem o timo e viajam com os septos da cápsula para a área entre o córtex e a medula, onde entram no próprio timo; ou, como alternativa, entre diretamente na cápsula.[3]
As veias do timo terminam na veia braquiocefálica esquerda, na veia torácica interna e nas veias inferiores da tireóide.[3] Às vezes, as veias terminam diretamente na veia cava superior.[3]
Os vasos linfáticos viajam apenas para longe do timo, acompanhando as artérias e veias. Eles drenam para os linfonodos braquiocefálicos, traqueobrônquicos e paraesternais.[3]
Os nervos que suprem o timo surgem do nervo vago e da cadeia simpática cervical.[3] Ramos dos nervos frênicos atingem a cápsula do timo, mas não entram no timo.[3]
Variação
[editar | editar código-fonte]Os dois lobos diferem ligeiramente em tamanho, com o lobo esquerdo geralmente mais alto que o direito. O tecido tímico pode ser encontrado espalhado na glândula ou ao redor dela e, ocasionalmente, dentro da tireóide.[3] O timo em crianças se estende variavelmente para cima, às vezes até a glândula tireóide.[3]
Desenvolvimento
[editar | editar código-fonte]
Os timócitos e o epitélio do timo têm diferentes origens de desenvolvimento.[5] O epitélio do timo se desenvolve primeiro, aparecendo como duas conseqüências, uma de cada lado, da terceira bolsa faríngea.[5] Às vezes, também envolve a quarta bolsa faríngea.[4] Estes se estendem para fora e para trás no mesoderma circundante e no mesênquima derivado da crista neural na frente da aorta ventral. Aqui, os timócitos e o epitélio se encontram e se juntam ao tecido conjuntivo. A abertura faríngea de cada divertículo é logo obliterada, mas o colo do balão persiste por algum tempo como cordão celular. Por uma proliferação adicional das células que revestem o frasco, formam-se gomos de células, que ficam cercadas e isoladas pelo mesoderma invasor.[7]
O epitélio forma lóbulos finos e se desenvolve como uma estrutura esponjosa. Durante esse estágio, os precursores hematopoiéticos da medula óssea migram para o timo.[5] O desenvolvimento normal depende da interação entre o epitélio e os timócitos hematopoiéticos. O iodo também é necessário para o desenvolvimento e atividade do timo.[8]
Involução
[editar | editar código-fonte]O timo continua a crescer após o nascimento, atingindo o tamanho máximo relativo na puberdade.[3] É mais ativo na vida fetal e neonatal.[9] Aumenta para 20 - 50 gramas na puberdade.[4] Em seguida, começa a diminuir em tamanho e atividade em um processo chamado involução tímica.[5] Após o primeiro ano de vida, a quantidade de linfócitos T produzidas começa a cair.[5] O tecido adiposo e o tecido conjuntivo preenchem uma parte do volume tímico.[3] Durante a involução, o timo diminui de tamanho e atividade.[5] As células adiposas estão presentes no nascimento, mas aumentam em tamanho e número acentuadamente após a puberdade, invadindo a glândula das paredes entre os lóbulos primeiro, depois para o córtex e a medula.[5] Esse processo continua na velhice, onde, seja com um microscópio ou com o olho humano, o timo pode ser difícil de detectar,[5] embora normalmente pesa de 5 a 15 gramas.[4]
A atrofia é devida ao aumento do nível circulante de hormônios sexuais, e a castração química ou física de um adulto resulta no aumento do tamanho e da atividade do timo.[10] Doenças graves ou infecção pelo vírus da imunodeficiência humana também podem resultar em involução.[4]
Função
[editar | editar código-fonte]
Maturação de células T
[editar | editar código-fonte]O timo facilita a maturação dos linfócitos T, uma parte importante do sistema imunológico que fornece imunidade mediada por células.[11] As células T começam como precursores hematopoiéticos da medula óssea e migram para o timo, onde são denominados timócitos. No timo, eles passam por um processo de maturação, que envolve garantir que as células reajam contra antígenos ("seleção positiva"), mas que não reajam contra antígenos encontrados no tecido corporal ("seleção negativa").[11] Uma vez maduras, as células T emigram do timo para fornecer funções vitais no sistema imunológico.[11][12]
Cada célula T possui um receptor de célula T distinto, adequado para uma substância específica, chamada antígeno.[12] A maioria dos receptores de células T se liga ao complexo principal de histocompatibilidade das células do corpo. O MHC apresenta um antígeno para o receptor de células T, que se torna ativo se corresponder ao receptor específico de células T.[12] Para ser adequadamente funcional, uma célula T madura precisa ser capaz de se ligar à molécula do MHC ("seleção positiva") e não reagir contra antígenos que são realmente dos tecidos do corpo ("seleção negativa").[12] A seleção positiva ocorre no córtex e a seleção negativa ocorre na medula do timo.[13] Após esse processo, as células T que sobreviveram deixam o timo, regulado pela esfingosina-1-fosfato.[13] Mais maturação ocorre na circulação periférica.[13] Parte disso ocorre devido aos hormônios e citocinas secretados pelo timo, incluindo timulina, timopoietina e timosinas.[5]
Seleção positiva
[editar | editar código-fonte]As células T têm receptores distintos de células T. Esses receptores distintos são formados pelo processo de rearranjo do gene de recombinação V(D)J estimulado pelos genes RAG1 e RAG20.[13] Esse processo é propenso a erros e alguns timócitos não conseguem produzir receptores funcionais de células T, enquanto outros timócitos produzem receptores de células T que são auto-reativos.[14] Se um receptor funcional de células T for formado, o timócito começará a expressar simultaneamente as proteínas da superfície celular CD4 e CD8.[13]
A sobrevivência e a natureza da célula T dependem então de sua interação com as células epiteliais do timo circundantes. Aqui, o receptor de células T interage com as moléculas de MHC na superfície das células epiteliais.[13] Uma célula T com um receptor que não reage, ou reage fracamente, morrerá por apoptose. Uma célula T que reage sobreviverá e proliferará.[13] Uma célula T madura expressa apenas CD4 ou CD8, mas não os dois.[12] Isso depende da força da ligação entre o TCR e o MHC classe 1 ou classe 2.[13] Um receptor de células T que se liga principalmente à classe I do MHC tende a produzir uma célula T CD8 positiva "citotóxica" madura; um receptor de célula T que se liga principalmente ao MHC classe II tende a produzir uma célula T CD4 positiva.[14]
Seleção negativa
[editar | editar código-fonte]As células T que atacam as proteínas do próprio corpo são eliminadas no timo, denominadas "seleção negativa".[12] As células epiteliais na medula e as células dendríticas no timo expressam as principais proteínas de outras partes do corpo.[13] O gene que estimula isso é o AIRE.[12][13] Os timócitos que reagem fortemente aos antígenos próprios não sobrevivem e morrem por apoptose.[12][13] Algumas células T CD4 positivas expostas a auto-antígenos persistem como células reguladoras T.[12]
Relevância clínica
[editar | editar código-fonte]Imunodeficiência
[editar | editar código-fonte]Como o timo é o órgão do desenvolvimento das células T, qualquer defeito congênito na gênese tímica ou um defeito no desenvolvimento do timócito pode levar a uma profunda deficiência de células T na doença primária da imunodeficiência.[9] A perda do timo em idade precoce por mutação genética (como na síndrome de Digeorge, síndrome de CHARGE ou um timo "nu" raro que causa ausência de pelos e do timo[15]) resulta em imunodeficiência grave e subseqüente alta suscetibilidade à infecção por vírus, protozoários e fungos.[9] Camundongos nus com a rara deficiência de "nus" como resultado da mutação FOXN1 são uma variedade de camundongos de pesquisa como modelo de deficiência de células T.[16]
A causa congênita mais comum de deficiência imunológica relacionada ao timo resulta da exclusão do cromossomo 22, chamada síndrome de DiGeorge.[9][15] Isso resulta em falha no desenvolvimento da terceira e quarta bolsas faríngeas, resultando em falha no desenvolvimento do timo e em vários outros problemas associados, como doenças cardíacas congênitas e anormalidades da boca (como fenda palatina e lábio leporino), falha no desenvolvimento das glândulas paratireóides e presença de uma fístula entre a traqueia e o esôfago.[9] Números muito baixos de células T circulantes são vistos.[9] A condição é diagnosticada por hibridização fluorescente in situ e tratada com transplante de timo.[15]
A imunodeficiência combinada grave (SCID) é um grupo de doenças genéticas congênitas raras que podem resultar em deficiências combinadas de células T, B e NK.[9] Essas síndromes são causadas por mutações que afetam a maturação das células progenitoras hematopoiéticas, que são precursoras das células B e T.[9] Vários defeitos genéticos podem causar SCID, incluindo perda de função do gene do receptor de IL-2 e mutação, resultando em deficiência da enzima adenina deaminase.[9]
Fisiologia
[editar | editar código-fonte]Em termos fisiológicos, o timo elabora várias substâncias: timosina alfa, timopoetina, timulina e o fator tímico circulante. A timosina mantém e promove a maturação de linfócitos e órgãos linfoides como o baço e linfonodos. Existe ainda uma outra substância, a timulina, que exerce função na placa motora (junção dos nervos com os músculos) e, portanto, nos estímulos neurais e periféricos, sendo considerada grande responsável por uma doença muscular chamada miastenia grave.
Órgão glandular endócrino transitório, próprio da infância, de que só restam vestígios adiposos aos 10-12 anos de idade. Encontra-se localizado na parte inferior do pescoço e superior do mediastino superior, formado por dois lobos alongados, compostos cada um deles por uma série de lóbulos dispostos em volta de um eixo ou cordão central do coração.
Cada lóbulo está dividido, por tabiques de tecido conjuntivo, em folículos formados por uma substância cortical e outra medular contendo linfócitos, leucócitos e corpúsculos de Hassal. O timo de animais foi empregue na terapêutica do bócio exoftálmico.
Imagens adicionais
[editar | editar código-fonte]-
 Timo de um feto
Timo de um feto -
 Na radiografia de tórax, o timo aparece como uma massa radiodensa (mais brilhante nesta imagem) pelo lobo superior do pulmão direito (esquerdo da imagem) da criança.
Na radiografia de tórax, o timo aparece como uma massa radiodensa (mais brilhante nesta imagem) pelo lobo superior do pulmão direito (esquerdo da imagem) da criança.


Referências
[editar | editar código-fonte]- Este artigo incorpora texto em domínio público da 20ª edição de Gray's Anatomy (1918)
- ↑ Infopédia. «timo | Dicionário Infopédia da Língua Portuguesa». Infopédia - Porto Editora. Consultado em 5 de junho de 2022
- ↑ a b c d e f g Standring, Susan; et al. (2008). Gray's anatomy: the anatomical basis of clinical practice 40 ed. Londres: Churchill Livingstone
- ↑ a b c d e f g h i j k l Standring, Susan; et al. (2016). Gray's anatomy: the anatomical basis of clinical practice 41 ed. Philadelphia: [s.n.] pp. 983–6
- ↑ a b c d e f g h i Chapter 13, Diseases of White Blood Cells, Lymph Nodes, Spleen, and Thymus: Thymus. Robbins and Cotran Pathologic Basis of Disease 9 ed. [S.l.: s.n.]
- ↑ a b c d e f g h i j k Young, Barbara; O'Dowd, Geraldine (2013). Wheater's functional histology: a text and colour atlas 6 ed. Philadephia: Elsevier. pp. 204–6
- ↑ Larsen, W (2001). Human Embryology 3 ed. [S.l.]: Elsevier. pp. 366–367
- ↑ Embryology. «Swiss embryology». Lymphatic tissue
- ↑ Venturi, S; Venturi, M (2009). Iodine, thymus, and immunity. [S.l.]: Nutrition. doi:10.1016%2Fj.nut.2009.06.002 Verifique
|doi=(ajuda) - ↑ a b c d e f g h i Ralston, Stuart H; et al. (2018). Davidson's principles and practice of medicine 23 ed. [S.l.]: Elsevier
- ↑ Sutherland, J. S. (2005). «Activation of thymic regeneration in mice and humans following androgen blockade». Journal of Immunology
- ↑ a b c Hall, John E. (2016). Guyton and Hall textbook of medical physiology 13 ed. Philadelphia: Elsevier. pp. 466–7
- ↑ a b c d e f g h i «Chapter 6. Diseases of the immune system. The normal immune system». Robbins and Cotran Pathologic Basis of Disease. [S.l.]: Elsevier
- ↑ a b c d e f g h i j k Hohl, Tobias M (2019). «6. Cell mediated defence against infection: Thymic selection of CD4+ and CD8+ T Cells». Mandell, Douglas, and Bennett's principles and practice of infectious diseases 9 ed. [S.l.]: Elsevier
- ↑ a b Miller, Jacques F. A. P (2011). «The golden anniversary of the thymus». Nature Reviews Immunology: 489–495. 2493 páginas
- ↑ a b c Kasper, Dennis; et al. (2015). Harrison's Principles of Internal Medicine 19 ed. [S.l.]: McGraw-Hill Professional
- ↑ Fox, James G (2006). The Mouse in Biomedical Research: Immunology. [S.l.]: Elsevier. 277 páginas