Chapter 2 Reading
Chapter 2 Reading
Download as pdf or txt
You might also like
- Leech Lab ReportDocument14 pagesLeech Lab Reportapi-339485098No ratings yet
- Control and Coordination - Study ModuleDocument50 pagesControl and Coordination - Study ModuleVivaanNo ratings yet
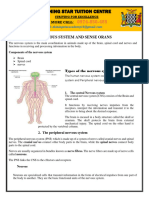
- 11.6. The Nervous System and Sense OrgansDocument12 pages11.6. The Nervous System and Sense OrgansIsaac SiameNo ratings yet
- Unit 5Document51 pagesUnit 5JAYANT CH (RA2111004010265)No ratings yet
- What Are NeuronsDocument16 pagesWhat Are NeuronsJaebeom's Best FriendNo ratings yet
- Ns-Elec-4 - Nervous SystemDocument14 pagesNs-Elec-4 - Nervous SystemONGLENGCO WILSON JAKE V.No ratings yet
- Neurons. The Building Blocks of Nervous SystemDocument3 pagesNeurons. The Building Blocks of Nervous SystemGinelQuenna De VillaNo ratings yet
- Nervous SystemDocument19 pagesNervous Systemmercaderlorenzo9No ratings yet
- Nervous SystemDocument9 pagesNervous SystemChristine Joanne CelendroNo ratings yet
- Invertebrate Nervous Systems: Thomas MathesonDocument6 pagesInvertebrate Nervous Systems: Thomas MathesonMax MoralesNo ratings yet
- Chap8 Nervous Transes-1-1Document8 pagesChap8 Nervous Transes-1-1y30ny30nNo ratings yet
- Ns-Elec-4 - Nervous SystemDocument14 pagesNs-Elec-4 - Nervous SystemONGLENGCO WILSON JAKE V.No ratings yet
- (Oct 1) Nervous-SystemDocument78 pages(Oct 1) Nervous-SystemBea Gualberto100% (1)
- MC1 REVIEWER (Nervous System) - MIDTERMSDocument7 pagesMC1 REVIEWER (Nervous System) - MIDTERMSFrancine Dominique CollantesNo ratings yet
- Central NervousDocument8 pagesCentral NervousDamian MaferiNo ratings yet
- (w14) Nervous Tissue and Nervous SystemDocument7 pages(w14) Nervous Tissue and Nervous Systemden mNo ratings yet
- Coordination Biology 1Document33 pagesCoordination Biology 1stevhobikhoNo ratings yet
- Unit 11 Nervous SystemDocument117 pagesUnit 11 Nervous SystemChandan Shah100% (1)
- Neurological SystemDocument21 pagesNeurological Systemapi-292000448No ratings yet
- Chapter 8 - Nervous ReviewerDocument18 pagesChapter 8 - Nervous Reviewerchristian anchetaNo ratings yet
- Nervous SystemDocument11 pagesNervous SystemYUAN FRANCIS SINGCULANNo ratings yet
- Topic 02A - The NeuronsDocument5 pagesTopic 02A - The NeuronsjmvengineerconsNo ratings yet
- Nervous System ResonanceDocument72 pagesNervous System ResonanceEkta ManglaniNo ratings yet
- Made Up of Neurons and Neuroglia Cells: Anatomical Subdivisions of NSDocument18 pagesMade Up of Neurons and Neuroglia Cells: Anatomical Subdivisions of NSJavorko PNo ratings yet
- Neural Contraol & Coordinaton - SNDocument7 pagesNeural Contraol & Coordinaton - SNPushpa DhruvNo ratings yet
- Physiology Handouts (Topic 16-26)Document35 pagesPhysiology Handouts (Topic 16-26)bc200411046No ratings yet
- Anatomy Lect. 5 1 Nervous System OutilnesDocument17 pagesAnatomy Lect. 5 1 Nervous System OutilnesolowolayemogoodnessNo ratings yet
- Lecture 3 The Nervous SystemDocument11 pagesLecture 3 The Nervous SystemJanvi MehtaNo ratings yet
- The Nervous SystemDocument8 pagesThe Nervous Systemsahiniahamed2No ratings yet
- Nervous System-Double AwardDocument4 pagesNervous System-Double AwardTumisang PitiroseNo ratings yet
- NervusDocument8 pagesNervusAndika Anjani AgustinNo ratings yet
- Excitable TissueDocument117 pagesExcitable Tissueur.yared21100% (1)
- Types Structure and Function of NeuronsDocument7 pagesTypes Structure and Function of Neuronsvishnuchaudhari322No ratings yet
- Chapter 14 - Control and Coordination in Living BeingsDocument9 pagesChapter 14 - Control and Coordination in Living Beingssanalaptop7No ratings yet
- Histology: Neuron Cells Types and StructureDocument7 pagesHistology: Neuron Cells Types and StructureAli HayderNo ratings yet
- Nervous System-WPS OfficeDocument36 pagesNervous System-WPS OfficeShubhendu Chattopadhyay100% (1)
- NeuroconductivityDocument16 pagesNeuroconductivitysandrajoshy06No ratings yet
- GenBio2 Lesson 1 Muscular and Nervous TissuesDocument5 pagesGenBio2 Lesson 1 Muscular and Nervous TissuesAnne BeatrizNo ratings yet
- Nervous Tissue - UpdatedDocument129 pagesNervous Tissue - UpdatedPojangNo ratings yet
- New Microsoft Word Document 1Document1 pageNew Microsoft Word Document 1api-385694647No ratings yet
- Neurobiology - Option - IBDocument49 pagesNeurobiology - Option - IBJarek OleszczynskiNo ratings yet
- The Nervous SystemDocument3 pagesThe Nervous SystemA. MagnoNo ratings yet
- Konsep Dasar Ilmu Biokimia Dan Biologi Molekuler Untuk Sistem SarafDocument31 pagesKonsep Dasar Ilmu Biokimia Dan Biologi Molekuler Untuk Sistem SarafdiandraNo ratings yet
- ICSE Class X Nervous System NotesDocument20 pagesICSE Class X Nervous System Notesprogrammer.ron.akNo ratings yet
- Coordination and ResponseDocument8 pagesCoordination and ResponseEnaya MajidNo ratings yet
- Physiology 2324Document13 pagesPhysiology 2324know more factsNo ratings yet
- Coordination and Response in Animals - 052037Document8 pagesCoordination and Response in Animals - 052037MosesNo ratings yet
- Nervous System and Common PathologiesDocument25 pagesNervous System and Common PathologiesgerardwilmotNo ratings yet
- Chapter 2 - Structure and Functions of CellsDocument7 pagesChapter 2 - Structure and Functions of Cellsmanilyn dacoNo ratings yet
- Handout For Gen Bio 2-2Document14 pagesHandout For Gen Bio 2-2haniNo ratings yet
- Anaphy Nervous SystemDocument6 pagesAnaphy Nervous SystemAndrea SaldivarNo ratings yet
- Nervous ControlDocument15 pagesNervous ControlzoeakatNo ratings yet
- IPG 9 - Nervous System ChangesDocument50 pagesIPG 9 - Nervous System ChangesdeviNo ratings yet
- Neurology Assignment 1Document9 pagesNeurology Assignment 1workupindia7No ratings yet
- Transes Nervous SystemDocument13 pagesTranses Nervous SystemAlther LorenNo ratings yet
- Nervous and Endocrine SystemDocument126 pagesNervous and Endocrine SystemNimra Bashir CheemaNo ratings yet
- ANAPHY - Nervous SystemDocument5 pagesANAPHY - Nervous SystemMA. ANDREA NICOLE DURANNo ratings yet
- Nervous SystemDocument23 pagesNervous Systemde_me3No ratings yet
- Biological Bases of BehaviorDocument8 pagesBiological Bases of BehaviormysterioushumaneNo ratings yet
- Structure of NeuronsDocument2 pagesStructure of NeuronsEnz JosephNo ratings yet
- Imaging Chapter 4 Physics of RadiographyDocument2 pagesImaging Chapter 4 Physics of RadiographyYousef Ahmad2No ratings yet
- Technical Communications 6Document7 pagesTechnical Communications 6Yousef Ahmad2No ratings yet
- Introduction To Medical Imaging SystemsDocument13 pagesIntroduction To Medical Imaging SystemsYousef Ahmad2No ratings yet
- Introduction To BiomechanicsDocument6 pagesIntroduction To BiomechanicsYousef Ahmad2No ratings yet
- Introduction To Biomedical EngineeringDocument2 pagesIntroduction To Biomedical EngineeringYousef Ahmad2No ratings yet
- 10 1126@science Aau3644Document14 pages10 1126@science Aau3644nicolasNo ratings yet
- Physiology Chap5 (Membrane Potentials & Action Potentials)Document6 pagesPhysiology Chap5 (Membrane Potentials & Action Potentials)Man Dejelo100% (1)
- Nervous Tissue NotesDocument3 pagesNervous Tissue NotesTulibas Karl Jade J - BSN 1ENo ratings yet
- Fundamentals of Nervous System and Nervous TissueDocument37 pagesFundamentals of Nervous System and Nervous Tissue차지형No ratings yet
- Nervous System: Team MembersDocument8 pagesNervous System: Team MembersnehaNo ratings yet
- 12.1 Coordination & Response, 12.2 Nervous System, 12.3 Neurones & ResponseDocument4 pages12.1 Coordination & Response, 12.2 Nervous System, 12.3 Neurones & ResponseHAJAR LENNo ratings yet
- Introduction To RMP ND APDocument78 pagesIntroduction To RMP ND APHariharanNo ratings yet
- 9 Answers To End-Of-Chapter QuestionsDocument7 pages9 Answers To End-Of-Chapter Questionsalyss ramdathNo ratings yet
- DNAwesome The Nervous System Part 2 Crash Course AP 9Document1 pageDNAwesome The Nervous System Part 2 Crash Course AP 9Clemcy Pearl MonisNo ratings yet
- Biolistrik: DR - Susy OliviaDocument97 pagesBiolistrik: DR - Susy OliviaHartomas BumiharjoNo ratings yet
- Full Download Test Bank For Foundations of Behavioral Neuroscience (Paper) 9/E 9th Edition Neil R. Carlson File PDF Free All ChapterDocument50 pagesFull Download Test Bank For Foundations of Behavioral Neuroscience (Paper) 9/E 9th Edition Neil R. Carlson File PDF Free All Chapterdomcsisolip10100% (11)
- Biological Psychology Kalat 12th Edition Test BankDocument33 pagesBiological Psychology Kalat 12th Edition Test BankTimothy Moultrie100% (39)
- Neuron Infographics by SlidesgoDocument35 pagesNeuron Infographics by SlidesgodainaleewallaceNo ratings yet
- Nervous Tissue.: Dr. Zahid Mehmood DPT, MS-NMPTDocument25 pagesNervous Tissue.: Dr. Zahid Mehmood DPT, MS-NMPTZahid Mehmood100% (1)
- Neuroglial Cells and NeuronsDocument16 pagesNeuroglial Cells and NeuronsMalliga SundareshanNo ratings yet
- Lecture One QuestionsDocument11 pagesLecture One QuestionsMagdy Mohamed Abdallah AbdelkaderNo ratings yet
- Properties of Nerve FibresDocument15 pagesProperties of Nerve Fibresdandasenasourabh1No ratings yet
- What Is The Difference Between Efferent and Afferent NervesDocument1 pageWhat Is The Difference Between Efferent and Afferent NervesranjuhiNo ratings yet
- Nerve Cells 2Document22 pagesNerve Cells 2Paulo Justin Tabangcora OropillaNo ratings yet
- The Cells of The Nervous System: Neurons and GliaDocument5 pagesThe Cells of The Nervous System: Neurons and GliaArcanus LorreynNo ratings yet
- Nervous System WSHDocument2 pagesNervous System WSHArdra AnilNo ratings yet
- Simple Reflex Arc Week 2Document4 pagesSimple Reflex Arc Week 2Ahmed KhaledNo ratings yet
- Functional Neuroanatomy Text and Atlas 2nd Edition by Adel AfifiDocument1,079 pagesFunctional Neuroanatomy Text and Atlas 2nd Edition by Adel AfifiPaola Andrea Martínez PérezNo ratings yet
- Test Bank For Seeleys Anatomy Physiology 12th by Vanputte DownloadDocument49 pagesTest Bank For Seeleys Anatomy Physiology 12th by Vanputte DownloadBenjaminWilsongaco100% (27)
- Chapter 5. Membrane Potential and Action PotentialDocument9 pagesChapter 5. Membrane Potential and Action Potentialjc timbNo ratings yet
- Biopsych Exam Questions PrelimDocument13 pagesBiopsych Exam Questions PrelimSharon Rose MedezNo ratings yet
- Nervous System MartiniDocument66 pagesNervous System MartiniHilma Halimatusy SyfaNo ratings yet
- 2nd Lecture - Synapse and Receptors - CNS Physiology (2nd Edition)Document16 pages2nd Lecture - Synapse and Receptors - CNS Physiology (2nd Edition)Mihaela Cenușe100% (1)
- Anggun Wening F - 1304617073 - Biolistrik Pada Sistem SarafDocument8 pagesAnggun Wening F - 1304617073 - Biolistrik Pada Sistem SarafRosa AmaliaNo ratings yet