Monoclonal Antibodies
Monoclonal Antibodies
Download as docx, pdf, or txt
You might also like
- 2023 Q3 HCPCSDocument58 pages2023 Q3 HCPCSRonadel RazonNo ratings yet
- The Clonogenic AssayDocument13 pagesThe Clonogenic AssayMohak Sahu100% (1)
- Abbott ABTDocument39 pagesAbbott ABTdunno2952No ratings yet
- Monoclonal AntibodyDocument8 pagesMonoclonal AntibodySadaf Sumdani NirvikNo ratings yet
- Bookchapter-Therapeutic Protein EngineeringDocument24 pagesBookchapter-Therapeutic Protein EngineeringDurgamadhab SwainNo ratings yet
- Azura Amid (Eds.) - Recombinant Enzymes - From Basic Science To Commercialization-Springer International Publishing (2015) PDFDocument191 pagesAzura Amid (Eds.) - Recombinant Enzymes - From Basic Science To Commercialization-Springer International Publishing (2015) PDFnurul qNo ratings yet
- NK-mediated Antibody-Dependent Cell-Mediated Cytotoxicity in Solid Tumors: Biological Evidence and Clinical PerspectivesDocument12 pagesNK-mediated Antibody-Dependent Cell-Mediated Cytotoxicity in Solid Tumors: Biological Evidence and Clinical PerspectivesTimelordNo ratings yet
- 2014 - Enzyme Assays PDFDocument15 pages2014 - Enzyme Assays PDFJason ParsonsNo ratings yet
- Affinity Chromatography - A Versatile Technique For Antibody PurificationDocument11 pagesAffinity Chromatography - A Versatile Technique For Antibody PurificationRaúl Sánchez OsunaNo ratings yet
- Cell Engineering and Cultivation of Chinese Hamster OvaryDocument8 pagesCell Engineering and Cultivation of Chinese Hamster OvaryDolce MaternitáNo ratings yet
- Affinity ChromatographyDocument368 pagesAffinity ChromatographySvarlgNo ratings yet
- An Imrpoved Yeast Transformation Method For The Generation of Very Large Human Antibody LibrariesDocument5 pagesAn Imrpoved Yeast Transformation Method For The Generation of Very Large Human Antibody Librariesbcn574835No ratings yet
- Angiogénesis Modelos, Moduladores y Aplicaciones ClínicasDocument552 pagesAngiogénesis Modelos, Moduladores y Aplicaciones ClínicasCarlos Sopán BenauteNo ratings yet
- Improving in Vivo Efficacy of Bioactive MoleculesDocument34 pagesImproving in Vivo Efficacy of Bioactive Moleculesירדן לויןNo ratings yet
- Therapeutic Targeting of Angiogenesis Molecular Pathways in Angiogenesis-Dependent DiseasesDocument11 pagesTherapeutic Targeting of Angiogenesis Molecular Pathways in Angiogenesis-Dependent Diseasesfelipe duarteNo ratings yet
- Production Recombinant Therapeutics in E. ColiDocument11 pagesProduction Recombinant Therapeutics in E. ColiStephany Arenas100% (2)
- The Biology of Apoptosis: Fouad Boulos, MD August 2010Document4 pagesThe Biology of Apoptosis: Fouad Boulos, MD August 2010ans11No ratings yet
- A New Runway For Purification of Messenger RNADocument10 pagesA New Runway For Purification of Messenger RNABruno AndreNo ratings yet
- Introduction of Life InsuranceDocument23 pagesIntroduction of Life InsuranceManwinder Singh GillNo ratings yet
- Protein Engineering and Its Applications in Food Industry 2017Document10 pagesProtein Engineering and Its Applications in Food Industry 2017SerkanNo ratings yet
- CHO Cell Line From Chinese Hamster Ovary: PropertiesDocument2 pagesCHO Cell Line From Chinese Hamster Ovary: PropertiesAhnaph RafinNo ratings yet
- Agar TechniqueDocument5 pagesAgar TechniqueMuhammad AjmalNo ratings yet
- Fluorescent Protein Bicistronic ExpressionDocument5 pagesFluorescent Protein Bicistronic ExpressionAlleleBiotechNo ratings yet
- A Cell-Based Model of Coagulation and The Role of Factor VIIa - Blood Review 2003Document5 pagesA Cell-Based Model of Coagulation and The Role of Factor VIIa - Blood Review 2003Oscar Echeverría OrellanaNo ratings yet
- Ig Structuer and FunctionsDocument34 pagesIg Structuer and FunctionsDentist Dina SamyNo ratings yet
- Tissue Factor and Breast CancerDocument9 pagesTissue Factor and Breast CancerBioscience and Bioengineering CommunicationsNo ratings yet
- Apatamers As TherapeuticsDocument14 pagesApatamers As TherapeuticsjazorelNo ratings yet
- Adaptive DesignDocument13 pagesAdaptive DesignChandan Kumar ChoubeyNo ratings yet
- Post Translational ModificationsDocument3 pagesPost Translational ModificationsIzzat RozaliNo ratings yet
- Medical Cell Biology: Xiaoying LiuDocument44 pagesMedical Cell Biology: Xiaoying LiuSayan KonarNo ratings yet
- The Klotho Gene and Soluble Klotho in Health and Disease From 1997 2018 A ReviewDocument13 pagesThe Klotho Gene and Soluble Klotho in Health and Disease From 1997 2018 A ReviewPaul HartingNo ratings yet
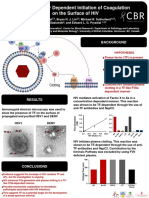
- Henry West - Tissue Factor Dependent Initiation of Coagulation On The Surface of HIVDocument1 pageHenry West - Tissue Factor Dependent Initiation of Coagulation On The Surface of HIVCBR UBCNo ratings yet
- CD MoleculesDocument25 pagesCD MoleculesunknownxemNo ratings yet
- Monoclonal AntibodiesDocument35 pagesMonoclonal Antibodiessam143143143143No ratings yet
- History of Medical BiotechnologyDocument18 pagesHistory of Medical BiotechnologyArisitica VinilescuNo ratings yet
- 235909Document15 pages235909kaviyabalakumarNo ratings yet
- 023.8 - Microcarrier Cell Culture PDFDocument171 pages023.8 - Microcarrier Cell Culture PDFSelvakapoorNo ratings yet
- Fluorescent Proteins and Their Applications in Imaging Living Cells and Tissues - by Chudakov Et Al., Physiol. Rev., 2010 1103-63Document62 pagesFluorescent Proteins and Their Applications in Imaging Living Cells and Tissues - by Chudakov Et Al., Physiol. Rev., 2010 1103-63Kassiani LiopetaNo ratings yet
- Applied Molecular Biology Beginning Laboratory ManualDocument60 pagesApplied Molecular Biology Beginning Laboratory ManualEmad ManniNo ratings yet
- Tissue Factor and Tissue Factor Pathway InhibitorDocument10 pagesTissue Factor and Tissue Factor Pathway Inhibitorfranciscrick69No ratings yet
- Natural ImmunityDocument34 pagesNatural ImmunitySelvaram SelvaNo ratings yet
- Protein Purification by Affinity Chromatography: January 2012Document21 pagesProtein Purification by Affinity Chromatography: January 2012Maria Villamizar GuerreroNo ratings yet
- Biology 310: Prokaryotic and Viral Genetics Plasmids: I. What Is A Plasmid?Document10 pagesBiology 310: Prokaryotic and Viral Genetics Plasmids: I. What Is A Plasmid?Syed NafeesNo ratings yet
- Antibody EngineeringDocument14 pagesAntibody EngineeringJholina Aure ReyesNo ratings yet
- M - 4 Basic Structure and Functions of ImmunoglobulinDocument10 pagesM - 4 Basic Structure and Functions of ImmunoglobulinDr. Tapan Kr. DuttaNo ratings yet
- Enzymatic Lysis and Disruption of Microbial CellsDocument5 pagesEnzymatic Lysis and Disruption of Microbial CellsSeptian JauhariansyahNo ratings yet
- Soft Agar Colony FormationDocument2 pagesSoft Agar Colony Formationharsha2733100% (1)
- 2018 Melincovici VegfDocument13 pages2018 Melincovici Vegfdusseldorf27No ratings yet
- Aptamers As Future DrugsDocument25 pagesAptamers As Future Drugspsc anandNo ratings yet
- Role of Chelated Mineral in Animal Production PerformanceDocument38 pagesRole of Chelated Mineral in Animal Production PerformanceBharathidhasan Selvarasu100% (1)
- Toxicological Profile For Zinc PDFDocument352 pagesToxicological Profile For Zinc PDFCrni Labud100% (1)
- Antibodies: Immunoglobulins - A Family of ProteinsDocument21 pagesAntibodies: Immunoglobulins - A Family of ProteinsMike zombieNo ratings yet
- GE AKTAdesignPurificationDocument160 pagesGE AKTAdesignPurificationWilla100% (1)
- Zinc in Health and DiseaseDocument33 pagesZinc in Health and DiseaseNikitha RafeekNo ratings yet
- Antibodies CHPT 4Document16 pagesAntibodies CHPT 4AmitAgarwal100% (1)
- Harmey - VEGF and CancerDocument185 pagesHarmey - VEGF and CancerBopiyudha bopiyudhaNo ratings yet
- Physicochemical Stability of The Antibody-Drug Conjugate PDFDocument8 pagesPhysicochemical Stability of The Antibody-Drug Conjugate PDFkmeriemNo ratings yet
- Biochemical Assessment of Vitamin DDocument24 pagesBiochemical Assessment of Vitamin DKarunya VkNo ratings yet
- Enzymes in Industry: Production and ApplicationsFrom EverandEnzymes in Industry: Production and ApplicationsWolfgang AehleNo ratings yet
- Chromatographic Methods in Clinical Chemistry and ToxicologyFrom EverandChromatographic Methods in Clinical Chemistry and ToxicologyRoger BertholfNo ratings yet
- Global Rheumatoid Arthritis Drugs MarketDocument2 pagesGlobal Rheumatoid Arthritis Drugs MarketiHealthcareAnalyst, Inc.No ratings yet
- (In English) 2020 Korea Innovative PHARMACEUTICAL CompanyDocument82 pages(In English) 2020 Korea Innovative PHARMACEUTICAL CompanyMariana HernandezNo ratings yet
- Feature: What's Fueling The Biotech Engine-2011 To 2012Document7 pagesFeature: What's Fueling The Biotech Engine-2011 To 2012jccntNo ratings yet
- 1.5 How A Drug Company Made $114 Billion by Gaming The U.SDocument7 pages1.5 How A Drug Company Made $114 Billion by Gaming The U.SUmay NihanNo ratings yet
- Abbott PowerPoint Slide Style GuideDocument22 pagesAbbott PowerPoint Slide Style GuideloicfrevilleNo ratings yet
- Best Practice & Research Clinical Rheumatology: Sergio SchwartzmanDocument12 pagesBest Practice & Research Clinical Rheumatology: Sergio SchwartzmanNobody 95No ratings yet
- SFA Therapeutics 12 Minute Overview Presentation 30 November 2021Document35 pagesSFA Therapeutics 12 Minute Overview Presentation 30 November 2021Ira SpectorNo ratings yet
- Studii CliniceDocument17 pagesStudii CliniceVlad MîndăianuNo ratings yet
- DR Adrian Jones - Seronegative SpondyloarthropathiesDocument38 pagesDR Adrian Jones - Seronegative SpondyloarthropathiesMaryam ShahbazNo ratings yet
- Aha Drug Pricing Study Report 01152019Document32 pagesAha Drug Pricing Study Report 01152019Mani DhamodharanNo ratings yet
- A Prospective Open-Label Clinical Trial of Adalimumab For The Treatment of Hidradenitis SuppurativaDocument6 pagesA Prospective Open-Label Clinical Trial of Adalimumab For The Treatment of Hidradenitis SuppurativapancholarpancholarNo ratings yet
- PsoriasisDocument11 pagesPsoriasisRizki SariWahyuniNo ratings yet
- India The Emerging Hub For Biologics and BiosimilarsDocument22 pagesIndia The Emerging Hub For Biologics and BiosimilarssanjitlNo ratings yet
- Horizon Scanning UKMI 2017Document57 pagesHorizon Scanning UKMI 2017piNo ratings yet
- Treatment of Erythrodermic Psoriasis With BiologicsDocument8 pagesTreatment of Erythrodermic Psoriasis With BiologicsJohnNo ratings yet
- How Chemistry Changed The World: Some Lighter Chemistry MilestonesDocument1 pageHow Chemistry Changed The World: Some Lighter Chemistry MilestonesEnrique RiverNo ratings yet
- Executive Summary: Leading Drug Safety Issues of 2012Document21 pagesExecutive Summary: Leading Drug Safety Issues of 2012aghniajolandaNo ratings yet
- Fact Sheet: BiologicsDocument4 pagesFact Sheet: BiologicsAldiarsoNo ratings yet
- Inflammation-Immunology FINALDocument30 pagesInflammation-Immunology FINALYash ThoratNo ratings yet
- Abott LaboratoriesDocument41 pagesAbott Laboratoriesnishant_gupta_22No ratings yet
- Commercial Considerations of Drug Discovery ProgramDocument549 pagesCommercial Considerations of Drug Discovery ProgramparinitaNo ratings yet
- Arthritis Juvenile Idiopathic ArthritisDocument6 pagesArthritis Juvenile Idiopathic Arthritismkyousuf100% (1)
- Patients and Nurses Preferences of Autoinjectors For RheumatDocument12 pagesPatients and Nurses Preferences of Autoinjectors For RheumatRashmi PardeshiNo ratings yet
- Rituxan RituximabDocument21 pagesRituxan RituximabMavisNo ratings yet
- Psoriasis: Psoriasis Type of PsoriasisDocument18 pagesPsoriasis: Psoriasis Type of PsoriasisDeepikaNo ratings yet
- Rheumatoid ArthritisDocument9 pagesRheumatoid ArthritisNader Smadi86% (7)
- AbbVie ReportDocument7 pagesAbbVie Reportakr200714No ratings yet
- PSB 419Document7 pagesPSB 419Erick PrasetyaNo ratings yet