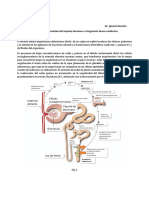
MerlydeArmas Rev - Cubanafarm.2006
MerlydeArmas Rev - Cubanafarm.2006
Descargar como pdf o txt
También podría gustarte
- Caso ClinicoDocumento5 páginasCaso ClinicoJosselin EnriquezAún no hay calificaciones
- AntiarritmicosDocumento11 páginasAntiarritmicosLaia Evelyn QYAún no hay calificaciones
- Caso Clinico EdumedDocumento3 páginasCaso Clinico EdumedNatali PalacioAún no hay calificaciones
- Sintesis Up 6Documento51 páginasSintesis Up 6Jennifer EspitiaAún no hay calificaciones
- Paper de FarmacologíaDocumento14 páginasPaper de FarmacologíafernandaAún no hay calificaciones
- Actualizaciones Recientes Sobre Nuevas Dianas Terapéuticas de Enfermedades CardiovascularesDocumento3 páginasActualizaciones Recientes Sobre Nuevas Dianas Terapéuticas de Enfermedades CardiovascularesMariangel PadillaAún no hay calificaciones
- SeminarioDocumento23 páginasSeminariovalentinaAún no hay calificaciones
- CardiotonicosDocumento4 páginasCardiotonicosDanny Chui ZodiaxAún no hay calificaciones
- CV Enfuac 2022Documento86 páginasCV Enfuac 2022Francisca AbarcaAún no hay calificaciones
- Choque CardiogenicoDocumento45 páginasChoque CardiogenicoGEOVANY CHIAún no hay calificaciones
- Módulo Dr. J. C. Crovetto. 2da. Parte. Farmacología en EnfermeríaDocumento10 páginasMódulo Dr. J. C. Crovetto. 2da. Parte. Farmacología en EnfermeríaelsaapaleAún no hay calificaciones
- CirculatorioDocumento19 páginasCirculatoriofabiola-Aún no hay calificaciones
- CardiotónicoDocumento10 páginasCardiotónicoCony XdAún no hay calificaciones
- Exposición Hipertensión Arterial CADocumento17 páginasExposición Hipertensión Arterial CAGaspar Ceballos ZuritaAún no hay calificaciones
- Farmacos AntihipertensivosDocumento50 páginasFarmacos Antihipertensivosandreina bestAún no hay calificaciones
- DFF - Efectos de La Astaxantina en La Funcion VascularDocumento281 páginasDFF - Efectos de La Astaxantina en La Funcion VascularKeithSix Yman Arrieta100% (1)
- Articulo Cientifico de Corazon AisladoDocumento10 páginasArticulo Cientifico de Corazon AisladoOscar Agustín JacoboAún no hay calificaciones
- Implementacion de La Tecnica en Organo AisladoDocumento6 páginasImplementacion de La Tecnica en Organo AisladoLuisito LaosAún no hay calificaciones
- Enrique Lopez Herrero Parte1 SabDocumento25 páginasEnrique Lopez Herrero Parte1 SabJEFFERSON MORALESAún no hay calificaciones
- FarmacosantiarritmicosDocumento42 páginasFarmacosantiarritmicosJesus de LaraAún no hay calificaciones
- Sistema CardiovascularDocumento13 páginasSistema Cardiovascularpaula vegaAún no hay calificaciones
- Informe Práctica 3 AntiarritmicosDocumento10 páginasInforme Práctica 3 AntiarritmicosBeto FernandezAún no hay calificaciones
- Sesión 8Documento5 páginasSesión 8evargasme1Aún no hay calificaciones
- Guia IccDocumento7 páginasGuia Iccdaniel.521826327Aún no hay calificaciones
- IVABRADINADocumento10 páginasIVABRADINAJaviera HidalgoAún no hay calificaciones
- Casos Clinicos HiperkalemiaDocumento44 páginasCasos Clinicos HiperkalemiahistoginoAún no hay calificaciones
- Tema 32. Parte 1. FarmaDocumento7 páginasTema 32. Parte 1. Farmamirrorsmith01Aún no hay calificaciones
- Acido Alfa Lipoico en La Neuropatía Diabética - NEW VERSIONDocumento67 páginasAcido Alfa Lipoico en La Neuropatía Diabética - NEW VERSIONJose Jesus ColonAún no hay calificaciones
- AntihipertermicosDocumento12 páginasAntihipertermicoskimyamileth117Aún no hay calificaciones
- Bloqueador de Los Canales de CalcioDocumento28 páginasBloqueador de Los Canales de CalcioHamer Meza Ubeda100% (1)
- Insuficiencia Cardiaca AgudaDocumento48 páginasInsuficiencia Cardiaca AgudaCarolinaAún no hay calificaciones
- Vasodilatadores FarmacologíaDocumento4 páginasVasodilatadores FarmacologíaAngelica RamirezAún no hay calificaciones
- 3..farmacologia Hipertension ArterialDocumento52 páginas3..farmacologia Hipertension ArterialINVERSIONES LUCIANAAún no hay calificaciones
- CASO CLÍNICO UPC Shock HipovolemicoDocumento5 páginasCASO CLÍNICO UPC Shock HipovolemicoRoma Cid GandulfoAún no hay calificaciones
- Presion Arterial, Fsiopatologia Hta 2016Documento63 páginasPresion Arterial, Fsiopatologia Hta 2016Herbert Martin Colan TorresAún no hay calificaciones
- HTA y SCADocumento23 páginasHTA y SCALuis Javier TarazonaAún no hay calificaciones
- Caso Clinico Grupo Fisiologia IDocumento13 páginasCaso Clinico Grupo Fisiologia IEDER ANTONIO DA COSTAAún no hay calificaciones
- AraiiDocumento6 páginasAraiiMoronta Ortiz Hector ManuelAún no hay calificaciones
- Bioquimica Caso ClinicDocumento7 páginasBioquimica Caso ClinicCatalina AlexandraAún no hay calificaciones
- Efectos Pleoitrooicos de CalciantagonistasDocumento8 páginasEfectos Pleoitrooicos de CalciantagonistasZabdi CastellanosAún no hay calificaciones
- Cap 112 PDFDocumento3 páginasCap 112 PDFadriana9mireya9viafaAún no hay calificaciones
- Electrolitos ABDocumento60 páginasElectrolitos ABRuben Eduardo GarciaAún no hay calificaciones
- Cardiología para El Estudiante PDFDocumento3 páginasCardiología para El Estudiante PDFLuisa Eduardo Sánchez CabezudoAún no hay calificaciones
- HiperpotasemiaDocumento2 páginasHiperpotasemiamonicafrezziniAún no hay calificaciones
- Clase 2Documento20 páginasClase 2Imef SumireAún no hay calificaciones
- Practica 5 AlbertDocumento13 páginasPractica 5 AlbertPUNTO CONTROL PUCARA PCPAún no hay calificaciones
- Antagonistas de Los Receptores de Angiotensina IIDocumento28 páginasAntagonistas de Los Receptores de Angiotensina IIacuacohauilaAún no hay calificaciones
- Fármacos y Corazón PDFDocumento17 páginasFármacos y Corazón PDFJonatan Aliaga RodriguezAún no hay calificaciones
- CAPITULO - 26 - DE - FARMACOLOGIA - IECA - Tamara - Docx Filename - UTF-8''CAPITULO 26 DE FARMACOLOGIA IECA TamaraDocumento6 páginasCAPITULO - 26 - DE - FARMACOLOGIA - IECA - Tamara - Docx Filename - UTF-8''CAPITULO 26 DE FARMACOLOGIA IECA TamaraAlejandra Perez RemisAún no hay calificaciones
- Farmacos Que Antagonizan Los Conductos de CalcioDocumento12 páginasFarmacos Que Antagonizan Los Conductos de CalcioOliverGlezAún no hay calificaciones
- ApuntesDocumento20 páginasApuntesJosé SalvadoAún no hay calificaciones
- Perfil CardiacoDocumento24 páginasPerfil CardiacoLuis Felipe OrtizAún no hay calificaciones
- El Agua de Aguas Profundas Modula La Presión Arterial y Exhibe Efectos Hipolipemiantes A Través de La Vía AMPK-ACCDocumento34 páginasEl Agua de Aguas Profundas Modula La Presión Arterial y Exhibe Efectos Hipolipemiantes A Través de La Vía AMPK-ACCgocardiseAún no hay calificaciones
- Guia 08 Farmacologia Grupo 1Documento5 páginasGuia 08 Farmacologia Grupo 1ALDER FRIEDERICH YACILA ESPINOZAAún no hay calificaciones
- Hipertension Arterial SistemicaDocumento34 páginasHipertension Arterial SistemicaHugo Robles Gómez100% (1)
- Inhibidores EcaDocumento38 páginasInhibidores EcaOscar Gonzalo Morales SilasAún no hay calificaciones
- Hipopotasemia e HiperpotasemiaDocumento43 páginasHipopotasemia e HiperpotasemiaROLANDOAún no hay calificaciones
- Guia de Estudio Arritmias, Grupo ADocumento7 páginasGuia de Estudio Arritmias, Grupo AJuan David FrancoAún no hay calificaciones
- Fundamentos Ekg & Síndrome Coronario Agudo PDFDocumento38 páginasFundamentos Ekg & Síndrome Coronario Agudo PDFAlejandra SuazzAún no hay calificaciones
- Examen Final Clinica Quirúrgica 1Documento21 páginasExamen Final Clinica Quirúrgica 1Guadalupe Lizeth A VAún no hay calificaciones
- Clasificación de Los Grupos FarmacológicosDocumento12 páginasClasificación de Los Grupos FarmacológicosyumalaislopezAún no hay calificaciones
- Hemorragia SubaracnoideaDocumento15 páginasHemorragia SubaracnoideaLeandro De JesusAún no hay calificaciones
- 1 Valoración de Signos VitalesDocumento30 páginas1 Valoración de Signos VitalesGeraldine Nicole Ramírez AlianteAún no hay calificaciones
- Accidente Cerebro VascularDocumento39 páginasAccidente Cerebro VascularJose Daniel Cardona Plata100% (1)
- Algo1. Similes Primer ParcialDocumento46 páginasAlgo1. Similes Primer ParcialGiovanna ForamiglioAún no hay calificaciones
- Resumen Fisio 2Documento83 páginasResumen Fisio 2Diego AbarcaAún no hay calificaciones
- Prueba de CoagulaciónDocumento24 páginasPrueba de CoagulaciónReyna LucioAún no hay calificaciones
- Factores Que Provocan Un Corazón Hipoeficaz: Punto 2 Punto 1Documento2 páginasFactores Que Provocan Un Corazón Hipoeficaz: Punto 2 Punto 1nikAún no hay calificaciones
- Artritis Reumatoide - Farm. ClinicaDocumento39 páginasArtritis Reumatoide - Farm. ClinicaRoxana Quincho SeguraAún no hay calificaciones
- Miocardiopatia Hipertrofica ObstructivaDocumento42 páginasMiocardiopatia Hipertrofica ObstructivaRobert Lozano PurizacaAún no hay calificaciones
- Modulo I HTADocumento26 páginasModulo I HTApalomazul007Aún no hay calificaciones
- Aparato RespiratorioDocumento10 páginasAparato Respiratoriodiana_altamirano12350% (2)
- ArteriosclerosisDocumento24 páginasArteriosclerosisValeria Gutiérrez100% (1)
- Cronofarmacologia GuíaDocumento7 páginasCronofarmacologia GuíaTamara Valdivia100% (3)
- Ruidos CardiacosDocumento40 páginasRuidos CardiacosMel Ruiz UAún no hay calificaciones
- Puntos Ventanas Al Cielo 1Documento5 páginasPuntos Ventanas Al Cielo 1Walter TaboadaAún no hay calificaciones
- Circulación SanguíneaDocumento6 páginasCirculación SanguíneaAlexander GomezAún no hay calificaciones
- Hernia Del Hiato EsofagicoDocumento12 páginasHernia Del Hiato EsofagicoJesus Diaz DelgadilloAún no hay calificaciones
- Anticoagulantes, Hemostáticos, Fibrinolíticos y Antiplaquetarios - CorrectoDocumento24 páginasAnticoagulantes, Hemostáticos, Fibrinolíticos y Antiplaquetarios - CorrectoGianella Zavala Gonzales100% (1)
- Sindromes Tiroideos Sin TXDocumento62 páginasSindromes Tiroideos Sin TXlady penelopeAún no hay calificaciones
- Anemias y PoliglobuliasDocumento3 páginasAnemias y PoliglobuliasSamuel RodríguezAún no hay calificaciones
- ENFERMEDAD ARTERIAL PERIFÉRICA - SabistonDocumento16 páginasENFERMEDAD ARTERIAL PERIFÉRICA - SabistonPablo BosaAún no hay calificaciones
- Manual de Metas Internacionales UrgenciasDocumento235 páginasManual de Metas Internacionales UrgenciasTavo Esc0% (1)
- DDJJ Cma 368368Documento4 páginasDDJJ Cma 368368federico leguizamonAún no hay calificaciones
- Resumen La CabergolinaDocumento2 páginasResumen La CabergolinaRocio Julcamoro TerronesAún no hay calificaciones
- Cirugía I y II Franco Bornancini (Resúmen)Documento48 páginasCirugía I y II Franco Bornancini (Resúmen)kayworkersAún no hay calificaciones
- Expo Neumotaponador FijacionDocumento17 páginasExpo Neumotaponador FijacionAndres LopezAún no hay calificaciones
- Anatomía III SucioDocumento29 páginasAnatomía III SucioNora El KhamlichiAún no hay calificaciones
- Aparatos para La Función de NutriciónDocumento21 páginasAparatos para La Función de NutriciónJulia BaliñoAún no hay calificaciones