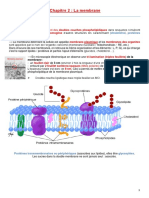
Chap - 3 - Les Molécules de Surface
Chap - 3 - Les Molécules de Surface
Télécharger au format docx, pdf ou txt
Vous aimerez peut-être aussi
- 05 Les Molecules D - AdhesionDocument4 pages05 Les Molecules D - AdhesionOns H'maiedPas encore d'évaluation
- Cours #1 - Jonction CellDocument96 pagesCours #1 - Jonction CellMzn JannahPas encore d'évaluation
- Cour TD de La Biologie CellulaireDocument13 pagesCour TD de La Biologie CellulairePiervie Guy Rosine BayidikilaPas encore d'évaluation
- Adherence 2017Document26 pagesAdherence 2017laarabPas encore d'évaluation
- CytologieDocument7 pagesCytologiedorissePas encore d'évaluation
- Cours de Biologie Cellulaire L3 BMCDocument16 pagesCours de Biologie Cellulaire L3 BMCIlyasse AitPas encore d'évaluation
- Adhérences Cellulaires 2019Document28 pagesAdhérences Cellulaires 2019imane.hniene123Pas encore d'évaluation
- Chap - 4 - Les Jonctions IntercellulairesDocument5 pagesChap - 4 - Les Jonctions IntercellulairesMoeanau GrangerPas encore d'évaluation
- Chapitre 2Document7 pagesChapitre 2dyhp2pswsrPas encore d'évaluation
- Cours Adhérence Cellulaire 1Document6 pagesCours Adhérence Cellulaire 1mohamed mkPas encore d'évaluation
- UE2 - Biologie Cellulaire - Molécule de Surface Et D - AdhérenceDocument19 pagesUE2 - Biologie Cellulaire - Molécule de Surface Et D - Adhérencesun-nee-chan9Pas encore d'évaluation
- Cellule Et EnvironnementDocument20 pagesCellule Et Environnementlamarisarah.pcPas encore d'évaluation
- COURS - 21-Larchitecture Cell Les JonctionsDocument63 pagesCOURS - 21-Larchitecture Cell Les JonctionsMeriem GhouarPas encore d'évaluation
- 07 - Adhérence CellulaireDocument6 pages07 - Adhérence CellulaireFairy MedPas encore d'évaluation
- 3ème Chapitre RésuméDocument4 pages3ème Chapitre Résumékawtherbz8Pas encore d'évaluation
- La Matrice Extracellulaire 2015-2016 PDFDocument3 pagesLa Matrice Extracellulaire 2015-2016 PDFAnonymous wVr908lBcK100% (2)
- Chapitre 5 Adherence CellulaireDocument43 pagesChapitre 5 Adherence CellulaireMeriem GhouarPas encore d'évaluation
- Jonctions Serres MSDocument6 pagesJonctions Serres MSJean-Loïc BauchetPas encore d'évaluation
- Chapitre 4 OGO Jonctions CellulairesDocument26 pagesChapitre 4 OGO Jonctions Cellulairesmohamadbaghdadu7Pas encore d'évaluation
- La MembraneDocument23 pagesLa MembraneFanny GrouxPas encore d'évaluation
- III - Membrane P (5) 2021-2022 (Adhésivité 1)Document128 pagesIII - Membrane P (5) 2021-2022 (Adhésivité 1)Ikramilla KaramillaPas encore d'évaluation
- Chapitre 3Document2 pagesChapitre 3dyhp2pswsrPas encore d'évaluation
- Les Protéines D'adhésion Et de Reconnaissance CellulaireDocument39 pagesLes Protéines D'adhésion Et de Reconnaissance CellulaireFatima BENHAFSSAPas encore d'évaluation
- Bio Cell 6Document5 pagesBio Cell 6Khyarhoum BrahimPas encore d'évaluation
- 13 - Molécules DadhérenceDocument24 pages13 - Molécules DadhérenceLea VernaldePas encore d'évaluation
- l12 Microbio Chap1Document9 pagesl12 Microbio Chap1Alhassane BahPas encore d'évaluation
- Molécules D'adhérence Cours L1Document3 pagesMolécules D'adhérence Cours L1safemindPas encore d'évaluation
- Epitheliums de RevetementDocument33 pagesEpitheliums de Revetementnihallydia24Pas encore d'évaluation
- Chap - 6 - Les Compartiments IntracellulairesDocument4 pagesChap - 6 - Les Compartiments IntracellulairesMoeanau GrangerPas encore d'évaluation
- Biologie Cellulaire Cours #2Document12 pagesBiologie Cellulaire Cours #2wafa saidiPas encore d'évaluation
- Chapitre 2 Tissu Conjonctif LAMDA 2021Document40 pagesChapitre 2 Tissu Conjonctif LAMDA 2021LàkàmoràEnAlgériePas encore d'évaluation
- Atlas en Couleur DhistologieDocument55 pagesAtlas en Couleur DhistologieDenisco NkemgnePas encore d'évaluation
- BIOCELL3Document89 pagesBIOCELL3samPas encore d'évaluation
- Synthèse La CelluleDocument6 pagesSynthèse La CelluleKamil LahbabiPas encore d'évaluation
- Cours PC Chapitre III M1 Biochimie Appliquée MOUHOUB F 1Document12 pagesCours PC Chapitre III M1 Biochimie Appliquée MOUHOUB F 1Assema SalmiPas encore d'évaluation
- Ch4. Relation Structure-Fonction (Cytosquelette Et Contraction Musculaire)Document15 pagesCh4. Relation Structure-Fonction (Cytosquelette Et Contraction Musculaire)lilia.douaouyaPas encore d'évaluation
- Les Matrices ExtracellulairesDocument18 pagesLes Matrices ExtracellulairesAmï NãPas encore d'évaluation
- Cours Biologie Des Organismes Séance 3Document36 pagesCours Biologie Des Organismes Séance 3ibovyclaraPas encore d'évaluation
- Fichier Produit 3806Document2 pagesFichier Produit 3806BOULAMA HASSANA MASSAWPas encore d'évaluation
- Technicien TerritorialDocument7 pagesTechnicien TerritorialPriscilliaArchenPas encore d'évaluation
- Adhérence Intercellulaire Et MatricielleDocument3 pagesAdhérence Intercellulaire Et Matricielleaptitudes-rappel0qPas encore d'évaluation
- Relation Structure Membranaire Et Fonction de La CelluleDocument22 pagesRelation Structure Membranaire Et Fonction de La CelluleliliPas encore d'évaluation
- 1 - La Polarité Et L'organisation CellulaireDocument7 pages1 - La Polarité Et L'organisation CellulairemissflorellePas encore d'évaluation
- Tissus ConjonctifsDocument28 pagesTissus Conjonctifssophia ezzaraiiPas encore d'évaluation
- Cours de Microbiologie 2eme Année BiologieDocument13 pagesCours de Microbiologie 2eme Année BiologieKàdî David100% (11)
- Biologie CellulaireDocument297 pagesBiologie Cellulaireesmorad53Pas encore d'évaluation
- Membrane PlasmiqueDocument17 pagesMembrane PlasmiqueSami SamirPas encore d'évaluation
- 05 Noyau-InterphasiqueDocument10 pages05 Noyau-InterphasiqueTttttPas encore d'évaluation
- 3 Chap 2 Cellule Procaryote PRDocument84 pages3 Chap 2 Cellule Procaryote PRTemtem HmPas encore d'évaluation
- Ultrastructure Cellule TabDocument3 pagesUltrastructure Cellule Tabwalid.chahmi.ordPas encore d'évaluation
- Biologie - Cellulaire - Cour KamelDocument15 pagesBiologie - Cellulaire - Cour Kamelrekik hibaPas encore d'évaluation
- Split Config - enDocument9 pagesSplit Config - envy876twp6cPas encore d'évaluation
- 12 - Le CytosqueletteDocument22 pages12 - Le CytosqueletteLea VernaldePas encore d'évaluation
- Exercices - BioCell - Lecon9 - Matrice Extrcellulaire-Jonctions CellulairesDocument6 pagesExercices - BioCell - Lecon9 - Matrice Extrcellulaire-Jonctions Cellulairestoungaramoussa025Pas encore d'évaluation
- La Membrane Plasmique: I. GénéralitésDocument17 pagesLa Membrane Plasmique: I. GénéralitésAisSa KeRrøùùmPas encore d'évaluation
- 1-Structure Et Archirecture de La Membrane Plasmique NeuronaleDocument8 pages1-Structure Et Archirecture de La Membrane Plasmique NeuronalebayaPas encore d'évaluation
- Goshen Histo GeneraleDocument13 pagesGoshen Histo Generalewjf6kvcm7sPas encore d'évaluation
- REF Cytologie PVDocument11 pagesREF Cytologie PVHelena HINTZER--FALCOPas encore d'évaluation
- Membranes cellulaires: Les Grands Articles d'UniversalisD'EverandMembranes cellulaires: Les Grands Articles d'UniversalisPas encore d'évaluation
- Épigénétique: Les Grands Articles d'UniversalisD'EverandÉpigénétique: Les Grands Articles d'UniversalisPas encore d'évaluation
- Chap - 6 - Les Compartiments IntracellulairesDocument4 pagesChap - 6 - Les Compartiments IntracellulairesMoeanau GrangerPas encore d'évaluation
- Chap - 5 - Le CytosqueletteDocument12 pagesChap - 5 - Le CytosqueletteMoeanau GrangerPas encore d'évaluation
- Chap - 1 - Organisation de La MB PlasmiqueDocument13 pagesChap - 1 - Organisation de La MB PlasmiqueMoeanau GrangerPas encore d'évaluation
- Chap - 4 - Les Jonctions IntercellulairesDocument5 pagesChap - 4 - Les Jonctions IntercellulairesMoeanau GrangerPas encore d'évaluation
- Chap - 2 - Les Phénomènes de TransportDocument5 pagesChap - 2 - Les Phénomènes de TransportMoeanau GrangerPas encore d'évaluation
- Chap - 1 - La CelluleDocument2 pagesChap - 1 - La CelluleMoeanau GrangerPas encore d'évaluation