Dinozaury
Dinozaury (Dinosauria) – zróżnicowany klad zauropsydów. Pojawiły się w triasie, około 237 milionów lat temu. Stanowiły dominującą grupę kręgowców lądowych od początku jury (około 201 milionów lat temu) przez 135 milionów lat, do końca kredy (66 milionów lat temu), kiedy wymieranie kredowe (wyznaczające granicę kreda-trzeciorzęd i koniec mezozoiku) doprowadziło do zniknięcia większości linii. Zapis kopalny wskazuje, że w jurze ptaki wyewoluowały z dinozaurów zaliczanych do teropodów, wobec czego przez literaturę przedmiotu wydaną w XXI wieku uznawane są za grupę dinozaurów[1]. Przeżywając katastrofę sprzed 66 milionów lat, dzięki swym potomkom stały się kontynuatorami linii dinozaurów po czasy współczesne[2].
| Dinosauria | |||
| Owen, 1842 | |||
| Okres istnienia: trias środkowy–dziś | |||
 Przedstawiciele różnych grup dinozaurów: na górze Gastonia burgei i Triceratops horridus, w środku wróbel zwyczajny i Stegosaurus stenops, na dole Edmontosaurus regalis i Apatosaurus louisae | |||
| Systematyka | |||
| Domena | |||
|---|---|---|---|
| Królestwo | |||
| Typ | |||
| Podtyp | |||
| Gromada | |||
| Podgromada | |||
| Infragromada | |||
| (bez rangi) | archozaury | ||
| (bez rangi) | Ornithodira | ||
| Nadrząd |
dinozaury | ||
| Rzędy | |||
|
| |||
Do ostatnich dziesięcioleci XX wieku wszystkie grupy dinozaurów uznawano za wymarłe. Jednakże zapis kopalny wskazuje, że ptaki, nazywane dziś w języku angielskim avian dinosaurs („ptasie dinozaury”)[3], są współczesnymi potomkami opierzonych dinozaurów[4]. Wyewoluowały one ze swych teropodzich przodków w okresie jury[5]. Wobec tego ptaki stanowią jedyną linię dinozaurów, która przetrwała zagładę[6].
Wobec powyższego do dinozaurów zalicza się również ptaki[7][8][9]. Z kolei dinozaury nie będące ptakami określa się mianem non-avian dinosaurs[10][11][12], tłumaczonym na język polski jako „nieptasie dinozaury”[13], pojawia się też określenie „nieptasie teropody”[14].
Dinozaury to z taksonomicznego, morfologicznego i ekologicznego punktu widzenia zróżnicowana grupa zwierząt. Do ptaków zaliczanych jest obecnie ponad 10 000 gatunków[15], co czyni je najbardziej zróżnicowaną grupą kręgowców z wyjątkiem okoniokształtnych[a][16]. Dzięki dowodom kopalnym do 2006 paleontolodzy wyróżnili ponad 500 rodzajów[17] i ponad tysiąc gatunków nieptasich dinozaurów[18], podczas gdy w 2014 liczba opisanych rodzajów wynosiła 943[19]. Dinozaury zamieszkiwały wszystkie kontynenty – na wszystkich bowiem znajdowane są ich szczątki kopalne, jak i występują współczesne gatunki[20]. Zaliczają się do nich zarówno roślinożercy, jak i mięsożercy. Pierwotnie cechowała je dwunożność, ale wiele wymarłych grup obejmowało formy czworonożne, a także poruszające się zarówno na dwóch, jak i na czterech łapach. Liczne grupy wykształciły wyszukane struktury, jak rogi czy kryzy. Niektóre taksony wymarłe wykształcały także takie modyfikacje szkieletu, jak kolce czy pancerz. Potwierdzono składanie jaj i budowę gniazd u wszystkich grup. O ile dzisiejsze ptaki osiągają zazwyczaj niewielkie rozmiary, co wiąże się z umiejętnością lotu, formy prehistoryczne osiągały często duże rozmiary. Największe zauropody mogły osiągać nawet 58 m długości przy wysokości 9,25 m[21]. Wizja nieptasich dinozaurów jako jednorodnie gigantycznych jest jednak nieprawdziwa, wynika z większej tendencji do zachowywania się dużych kości. Wiele nieptasich dinozaurów było niewielkich rozmiarów, jak mierzący około 50 cm długości Xixianykus.
Nazwa dinozaury oznacza „straszne jaszczurki”, dlatego też może wprowadzać w błąd – dinozaury nie były jaszczurkami, ani nie były z nimi blisko spokrewnione. Reprezentują natomiast oddzielną grupę „gadów”, oprócz ptaków ich najbliższymi współczesnymi krewniakami są krokodyle. Jak wiele wymarłych form, nie wykazują one cech tradycyjnie uznawanych za „gadzie”, jak zmiennocieplność czy poruszanie się na kończynach odwiedzionych na boki. Co więcej, wiele grup prehistorycznych zwierząt, takich jak mozazaury, ichtiozaury, pterozaury, plezjozaury, a nawet Dimetrodon, popularnie uznawanych za dinozaury, w rzeczywistości nimi nie są. Przez pierwszą połowę XX wieku, zanim uznano ptaki za dinozaury, większość społeczności naukowej uznawała dinozaury za powolne i zmiennocieplne. Zmiana nastąpiła w latach 60. (odrodzenie dinozaurów). Stwierdzono, że wszystkie dinozaury, w tym mezozoiczne, prowadziły aktywny tryb życia i cechowały się szybkim metabolizmem, tak jak dzisiejsi ich przedstawiciele; wykazywały też liczne adaptacje umożliwiające interakcje społeczne.
Odkąd naukowcy zaczęli badać i opisywać pierwsze skamieliny nieptasich dinozaurów w pierwszej połowie XIX wieku, zmontowane szkielety nieptasich dinozaurów stały się głównymi atrakcjami muzeów wielu krajów, a dinozaury stały się częścią kultury. Wielkie rozmiary niektórych grup, jak również niekiedy przerażająca i fantastyczna natura, zapewniły tym stworzeniom regularną obecność w bestsellerach książkowych i filmowych, jak Zaginiony świat Arthura Conana Doyle’a czy Park Jurajski Michaela Crichtona. Trwały entuzjazm opinii publicznej dla tych zwierząt doprowadził do znacznego wsparcia finansowego dla poświęconej im dziedziny nauki. Media regularnie donoszą o nowych odkryciach.
Etymologia
edytujTakson Dinosauria został formalnie nazwany w 1842 przez paleontologa Richarda Owena, który użył tego terminu do określenia „odrębnego plemienia lub podrzędu gadów Sauria”[22]. Nazwa pochodzi od greckich słów δεινός (deinos, co znaczy „straszny”, „potężny”, „wzbudzający lęk”) oraz σαῦρος (sauros, oznaczającego „jaszczurkę” lub „gada”)[22][23]. Choć ta nazwa taksonomiczna bywała często interpretowana w odniesieniu do dinozaurzych zębów, pazurów i innych wzbudzających strach cech, Owen miał zamiar podkreślić głównie rozmiary i majestat tych stworzeń[24].
Definicja filogenetyczna
edytuj
Dinozaury definiuje się zazwyczaj jako grupę obejmującą Triceratops, Neornithes, ich ostatniego wspólnego przodka (MRCA) i wszystkich jego potomków[25]. Zasugerowano również definicję obejmującą ostatniego wspólnego przodka megalozaura i iguanodonta, a także wszystkich jego potomków, ponieważ wymienione dwa rodzaje należą do trzech wymienionych przez Owena, gdy ustanawiał Dinosauria[26]. Obie wskazują dokładnie ten sam zbiór zwierząt: „Dinosauria = Ornithischia + Saurischia”, obejmują więc zazwyczaj dwunożne mięsożerne teropody, do których zaliczają się też ptaki, czworonożne opancerzone ankylozaury, czworonożne stegozaury o płytach na grzbiecie, czworonożne roślinożerne ceratopsy o kostnych kryzach, dwu- lub czworonożne ornitopody, a prawdopodobnie i wielkie roślinożerne zauropodomorfy o długich szyjach i ogonach.

Wielu paleontologów podnosi, że punkt, w którym rozdzieliły się linie teropodów i zauropodomorfów, może wyłączać zauropodomorfy zarówno z definicji dinozaurów, jak i gadziomiednicznych. By uniknąć niestabilności, dinozaury można zdefiniować bardziej konserwatywnie, uwzględniając trzy główne linie: Triceratops horridus + Saltasaurus loricatus + Passer domesticus, co oznacza ich ostatniego wspólnego przodka wraz ze wszystkimi jego potomkami. Ta bezpieczniejsza definicja może zostać wyrażona jako „Dinosauria = Ornithischia + Sauropodomorpha + Theropoda”[27]. Baron, Norman i Barrett (2017) zdefiniowali Dinosauria w podobny sposób: jako najmniejszy klad, do którego należą Passer domesticus, Triceratops horridus i Diplodocus carnegii[28]. Langer i współpracownicy (2020) zdefiniowali Dinosauria jako najmniejszy klad obejmujący Iguanodon bernissartensis, Megalosaurus bucklandii i Cetiosaurus oxoniensis[29].
Opis ogólny
edytujUżywając jednej z powyższych definicji, dinozaury można ogólnie opisać jako archozaury z kończynami ustawionymi pionowo pod tułowiem[30]. Wiele prehistorycznych grup zwierząt popularnie łączy się z dinozaurami: ichtiozaury, mozazaury, plezjozaury, pterozaury, nawet Dimetrodon się wśród nich pojawia, jednak formy te nie są dinozaurami. Nie cechują się one wyprostowanymi kończynami charakterystycznymi dla dinozaurów[31]. Dinosauria stanowiły dominującą grupę kręgowców lądowych w mezozoiku, zwłaszcza w okresach jurajskim i kredowym. Inne grupy zwierząt były ograniczone pod względem zajmowanej niszy i przybieranych rozmiarów. Przykładowo ssaki rzadko przekraczały rozmiarami kota, były głównie mięsożercami wielkości współczesnych gryzoni polującymi na niewielką zdobycz[32].

Dinosauria były i są niezwykle zróżnicowaną grupą zwierząt. W publikacji z 2006 Steve Wang i Peter Dodson szacują całkowitą liczbę rodzajów nieptasich dinozaurów na około 1850. Do tego czasu opisanych zostało 527 rodzajów, tak więc wedle autorów 71% rodzajów nieptasich dinozaurów było wtedy nieznanych nauce[17]. W 2011 uznawano ponad 850 rodzajów[33]. Wcześniejsza publikacja przewiduje, że istniało około 3400 rodzajów dinozaurów, z których wiele nie przetrwało w zapisie kopalnym[34]. Do września 2008 nazwano 1047 różnych gatunków[18]. Wang i Dodson (2006) prognozują, że w przeciągu następnych 60–100 lat poznane zostanie ¾ wszystkich rodzajów nieptasich dinozaurów, a w czasie 100–140 lat – 90%[17].
Niektóre żywiły się pędami roślin, nasionami, inne mięsem, rybami, owadami, występowała wśród nich też wszystkożerność. Ich przodkowie poruszali się na dwóch kończynach. Niektóre, jak Ammosaurus czy Iguanodon, mogły przemieszczać się na dwóch bądź czterech łapach. Częste były też różne modyfikacje czaszki, jak rogi czy kryza. Zdarzał się też kostny pancerz. Wiele także mezozoicznych form osiągało wielkość człowieka lub mniejszą. Współczesne ptaki też zazwyczaj nie są duże. Dinozaury zajmują obecnie wszystkie kontynenty. Ich skamieniałości wskazują, że również w przeszłości cechowały się globalnym zasięgiem, chociażby w epoce jury wczesnej[20]. Dzisiaj zajmują większość nadających się siedlisk, zarówno lądowych, jak i morskich. Istnieją dowody na to, że niektóre nieptasie dinozaury, jak mikroraptor, potrafiły latać bądź przynajmniej szybować, a inne, jak spinozaury, wiodły częściowo wodny tryb życia[35].
Synapomorfie
edytujWspółczesne odkrycia sprawiły, że podanie uniwersalnej listy wyróżniających się cech dinozaurów, cechującej się pełną zgody specjalistów, jest bardzo trudne. Prawie wszystkie odkryte do tej pory dinozaury dzielą pewne modyfikacje archozaurzego szkieletu przodków. Choć pewne późniejsze grupy zmodyfikowały również te cechy, są one uznawane za typowe dla Dinosauria; posiadali je najwcześniejsi przedstawiciele i przekazali je swym potomkom. Te wspólne cechy danego taksonu określa się mianem synapomorfii.
Szczegółowa ocena powiązań archozaurów wykonana przez S. Nesbitta[36] potwierdziła lub znalazła 12 pewnych synapomorfii (część z nich znano już wcześniej)
- dół podskroniowy w czaszce naprzeciw otworu podskroniowego
- epipophyses w proksymalnych kręgach szyjnych (nie licząc atlasa i obrotnika)
- koniuszek grzebienia naramienno-piersiowego (miejsce przyczepu mięśni naramienno-piersiowych) położony na co najmniej 30% dolnej długości kości ramiennej
- kość promieniowa nie osiąga 80% długości kości ramiennej
- czwarty krętarz (wypukłość w miejscu przyczepu mięśni musculus caudofemoralis) na kości udowej jest ostro obrzeżony
- asymetryczny krętarz czwarty posiada brzeg dystalny tworzący strony kąt z trzonem
- proksymalna powierzchnia stawowa dla strzałki na kościach skokowej i piętowej zajmuje mniej niż 30% poprzecznego wymiaru tych kości
- kość bocznopotyliczna leżąca na tyle czaszki, nie spotyka się z kością jednoimienną w płaszczyźnie pośrodkowej jamy wewnątrzczaszkowej
- proksymalne powierzchnie stawowe kości kulszowej dla kości biodowej i łonowej rozdzielone dużą wklęsłą powierzchnią
- cnemial crest piszczeli, czyli grzebień na jej przedniej powierzchni, biegnie przednio-bocznie, a więc zakrzywia się ku przodowi i zewnętrznej stronie kości
- wydatny grzebień zorientowany proksymalno-dystalnie na tylnej powierzchni dalszego końca piszczeli
Nesbitt znalazł wiele dalszych potencjalnych synapomorfii, nie uznał zaś licznych synapomorfii zasugerowanych wcześniej. Niektóre z nich są bowiem obecne także u Silesauridae, wedle Nesbitta grupy siostrzanej dinozaurów. Zaliczają się doń duży przedni krętarz, kości śródstopia od II do IV prawie równej długości, redukcja kontaktu między kośćmi łonową i kulszową, obecność cnemial crest na piszczeli i wznoszący się wyrostek na kości skokowej[25] i wiele innych.


Dinozaury dzielą również różnorodne inne cechy szkieletu. Jednakże często spotyka się je i u innych grup archozaurów lub też nie występują u wszystkich wczesnych dinozaurów, dlatego nie można określić ich synapomorfiami. Dla przykładu dinozaury jako diapsydy miały pierwotnie dwie pary otworów skroniowych w czaszce (tzw. okna skroniowe leżące za oczami); jako zaś archozaury (grupa diapsydów) posiadają dodatkowe otwory: przedoczodołowy i w żuchwie[37]. Co więcej, kilka cech uznanych niegdyś za synapomorfie dinozaurów okazało się być starszymi od tej grupy lub też nie występowały u najwcześniejszych jej przedstawicieli, a rozwinęły się niezależnie u różnych grup dinozaurów. Wymienia się tutaj wydłużoną łopatkę, kość krzyżową złożoną z co najmniej trzech zrośniętych kręgów (3 występują u niektórych innych archozaurów, ale tylko 2 ma herrerazaur)[25] i panewkę stawu biodrowego z otworem w środku powierzchni wewnętrznej (zamknięta panewka występuje np. u saturnalii[38][39], wczesnego zauropodomorfa z Argentyny[40]). Inna trudność w identyfikacji wyróżniających dinozaury cech wynika z ubogiej wiedzy dotyczącej wczesnych dinozaurów i innych archozaurów oraz łączących je podobieństw w różnych aspektach. Zwierzęta te często były w literaturze źle klasyfikowane[41].
Dinosauria różnią się ułożeniem kończyn od większości gadów[42][43][44]. Kończyny czworonogów powstały z rybich płetw. Odziedziczyły po nich charakterystyczne ustawienie – w kierunku bocznym, prostopadle do głównej osi ciała. Jednakże chodzenie wiąże się z większym wydatkiem energetyczny niż pływanie, co więcej, kończyny rozstawione szeroko na boki jeszcze ten wydatek zwiększają. Tymczasem ułożenie kończyn prosto pod tułowiem zmniejsza pracę potrzebną na podnoszenie ciała. Kończyny ustawione częściowo pod tułowiem rozwinęły się u gadów innych niż dinozaury. Obecnie spotyka się je u krokodyli. U dinozaurów, jak już wspomniano, kończyny ustawione są całkowicie pod ciałem. Ich staw biodrowy jest też bardziej okrągły niż u przodków. Jednakże kość udowa nie ulegała rotacji, ruch w stawie odbywał się w dwóch wymiarach. Działająca na panewkę siła kieruje się w takim wypadku do góry, a nie przyśrodkowo. Przekłada się to na lżejszą budowę przyśrodkowej części panewki. Potwierdza to zapis kopalny – obserwuje się otwarte panewki nieptasich dinozaurów (leżąca w tym miejscu za życia chrząstka nie zachowuje się w skamieniałościach)[42].
Postawa na wyprostowanych łapach pozwoliła uniezależnić oddychanie od poruszania się[42], dzięki czemu ułatwiła wczesnym dinozaurom wymianę gazową podczas przemieszczania się, co pozwoliło osiągnąć wytrzymałość i poziom aktywności przewyższający poruszające się „pełzającym krokiem” gady[45]. Wyprostowane kończyny, potrafiące unieść większy ciężar[42], wsparły także prawdopodobnie ewolucję wielkich rozmiarów poprzez redukcję naprężeń zginających kończyn[46]. Niektóre niezaliczane do dinozaurów archozaury, jak rauizuchy, także dysponowały wyprostowanymi łapami, ale osiągnęły je dzięki innej budowie stawu biodrowego[46].
Opisane położenie kończyn pozwoliło również na dwunożność. Nieptasie dinozaury poruszające się na dwóch nogach cechuje bardziej cylindryczna głowa kości udowej. Na bocznej powierzchni szyjki znajdowało się wybrzuszenie przypominające z wyglądu krętarz większy, pokryte za życia chrząstką. Mogła się tam znajdować kaletka stawowa[42].
Historia ewolucji
edytujPoczątki
edytuj

Dinozaury wyewoluowały z ich archozaurzych przodków około 230 milionów lat temu w epoce środkowego bądź późnego triasu, mniej więcej 20 milionów lat po wymieraniu permskim, które wyeliminowało według szacunków 95% życia na Ziemi[47][48]. Datowanie radiometryczne formacji geologicznych zawierających skamieliny wczesnego dinozaura z rodzaju Eoraptor umieszcza go właśnie w tym czasie. Uważa się, że eoraptor przypomina wspólnego przodka wszystkich dinozaurów, co podaje Sereno[49]. Jeśli to prawda, sugeruje to, że pierwsze dinozaury były niewielkimi, dwunożnymi drapieżnikami[50]. Jednakże pogląd widzący w eoraptorze drapieżnika traci na aktualności. Uważa się go raczej za wszystkożercę, jeśli nie roślinożercę, a istnieje nawet pogląd umieszczający go wśród bazalnych zauropodomorfów[51]. Odkrycie prymitywnych, przypominających dinozaury przedstawicieli Ornithodira, jak argentyńskie Marasuchus i Lagerpeton ze środkowego triasu, wspiera ten pogląd. Analiza skamieniałości wskazuje, że byli to niewielcy drapieżnicy poruszający się dwunożnie. Dinozaury mogły jednak pojawić się już 243 miliony lat temu, czego dowodziłyby skamieliny Nyasasaurus z tego czasu. Jednakże znalezione szczątki tego zwierzęcia są zbyt fragmentaryczne, by można było orzec na pewno, czy jest to dinozaur, czy też może stanowi ich najbliższą grupę zewnętrzną[52]
Gdy pojawiły się dinozaury, siedliska lądowe zajmowały różnorodne archozauromorfy i terapsydy, jak aetozaury, cynodonty, dicynodonty, Ornithosuchidae, rauizuchy i rynchozaury. Większość z nich wymarła w triasie podczas jednego bądź dwu wymierań. Najpierw, około 215 milionów lat temu, uległy wymarciu różne bazalne archozauromorfy, w tym Prolacertiformes. Następnie około 200 milionów lat temu odbyło się wymieranie triasowe, które zmiotło z powierzchni Ziemi większość grup wczesnych archozaurów, jak aetozaury, Ornithosuchidae, fitozaury[25] (wedle nowszych poglądów uznawane za takson siostrzany archozaurów[53]) i Rauisuchia, a ponadto prawdopodobnie także dicynodonty. Spośród fauny lądowej przetrwały krokodylomorfy, dinozaury, ssaki, pterozaury i żółwie[25]. Pojedyncza linia dinozaurów uległa radiacji adaptacyjnej w triase późnym i jurze wczesnej (230-190 milionów lat temu), rozdzielając się na wiele osobnych linii ewolucyjnych, wedle długo akceptowanego poglądu wypierając dawniejsze formy z ich nisz ewolucyjnych, wedle nowszego zaś po prostu zajmując nisze pozostawione po wymieraniu na przełomie karniku i noryku[54].
Ewolucja i paleobiogeografia
edytujEwolucja dinozaurów po okresie triasu odzwierciedla zmiany w roślinności i położenia kontynentów. W późnym triasie i wczesnej jurze były one połączone w pojedynczy superkontynent Pangeę. Zamieszkiwała go ogólnoświatowa fauna złożona głównie z mięsożernych celofyzoidów i roślinożernych wczesnych zauropodomorfów[55]. Rośliny nagonasienne, zwłaszcza zaś iglaste, stanowiące potencjalne źródło pożywienia, uległy w późnym triasie radiacji. Wczesne zauropodomorfy nie dysponowały jeszcze wysublimowanymi mechanizmami rozdrabniania pokarmu w jamie ustnej, wobec czego musiały wykorzystywać inne sposoby rozdrabniania go w dalszych odcinkach przewodu pokarmowego[56]. Ogólne podobieństwo faun dinozaurów trwało do środkowej, a nawet późnej jury, kiedy większość terenów zamieszkiwały takie drapieżniki, jak ceratozaury, spinozaury i karnozaury oraz roślinożercy w postaci ptasiomiednicznych stegozaurów i olbrzymich zauropodów. Za przykład wystarczyć może północnoamerykańska formacja Morrison i Tendaguru z Tanzanii w Afryce. Pewne różnice wykazują dinozaury z terenów Chin. Żyły tam wyspecjalizowane teropody sinraptory (bliscy krewni allozaurów[57]) i niezwykłe długoszyje zauropody, jak mamenchizaur[55]. Prozauropody już wyginęły, coraz częstsze stawały się ankylozaury i ornitopody. Do najpopularniejszych roślin należały iglaste i Pteridophyta. Zauropody, jak wcześniejsze prozauropody, nie rozdrabniały pokarmu w jamie ustnej, ale ptasiomiedniczne wykształciły różnorodne sposoby rozdrabniania pokarmu już w jamie ustnej. Miały prawdopodobnie narządy przypominające policzki do trzymania pokarmu w jamie ustnej, miażdżyły pokarm ruchami szczęki i żuchwy[56]. Innym ważnym wydarzeniem ewolucyjnym jury było pojawienie się ptaków, które wykształciły się z celurozaurów z grupy maniraptorów[58].
We wczesnej kredzie w związku z rozpadem superkontynentu Pangei dinozaury zaczęły wykazywać znaczne zróżnicowanie na różnych masach lądowych. Najwcześniej rozprzestrzeniły się ankylozaury, iguanodonty i brachiozaury, zajmując Europę, Amerykę Północną i północ Afryki. Następnie uzupełniły je bądź wyparły w Afryce olbrzymie spinozaurydy i karcharodontozaurydy wśród teropodów, rebbachizaurydy i tytanozaury wśród zauropodów, spotykane także w Ameryce Północnej. W Azji stały się popularne celurozaury z grupy maniraptorów, w tym dromeozaurydy, troodonty i owiraptorozaury, ważnymi roślinożercami stały się ankylozaurydy i wczesne ceratopsy, jak psitakozaur. Australia była domem pierwotnych ornitopodów, zwierząt zwanych hipsylofodontami (obecnie, dawniej zaliczane tam rodzaje leżą w różnych miejscach drzewa ornitopodów[59]) i iguanodontów[55]. Stegozaury zaczęły wymierać w późnej kredzie wczesnej bądź we wczesnej kredzie późnej. Największa zmianę kredy wczesnej, która uległa jeszcze zwiększeniu w epoce kredy późnej, polegała na ewolucji roślin kwiatowych. W tym samym czasie kilka grup dinozaurów roślinożernych wykształciło bardziej wysublimowane przystosowania do rozdrabniania pokarmu w jamie ustnej. Ceratopsy rozwinęły metodę odkrajania zębami zgrupowanymi w bateriach, iguanodonty zaś miażdżyły pokarm bateriami zębowymi, czego ekstremalnym przykładem są hadrozaury[56]. Baterie zębowe pojawiły się również u niektórych zauropodów. Najlepszy przykład stanowi tutaj nigerzaur z rodziny rebbachizaurów[60].
Wyróżnia się trzy główne fauny dinozaurów kredy późnej. Na północnych kontynentach dzisiejszej Ameryki Północnej i Azji głównymi teropodami były tyranozaury i wiele różnych mniejszych maniraptorów. Roślinożercy zaliczali się głównie do ptasiomiednicznych. Wymienia się tu hadrozaury, ceratopsy, ankylozaury i pachycefalozaury. Na kontynentach południowych powstałych z rozpadu Gondwany najczęstszymi teropodami były abelizaury, roślinożercami zaś tytanozaury. W końcu w Europie dominowały teropody dromeozaury, rabdodony (z grupy iguanodontów), opancerzone nodozaury i zauropody tytanozaury[55]. Znacznie rozprzestrzeniały się rośliny kwiatowe[56], a pod koniec kredy pojawiły się pierwsze trawy[61]. Miażdżące pokarm kaczodziobe i ścinające go ceratopsy uległy olbrzymiemu zróżnicowaniu w Ameryce Północnej i Azji. Teropody także ulegały radiacji, powstawały formy wszystkożerne i roślinożerne: pospolite stały się ornitomimozaury i terizinozaury[56].
Koniec okresu kredowego wyznacza wymieranie, które wydarzyło się 66 milionów lat temu. Spowodowało ono zniknięcie z powierzchni Ziemi wszystkich dinozaurów z wyjątkiem ptaków Neornithes. Przeżyły też niektóre inne grupy diapsydów, jak krokodyle, żółwie, jaszczurki, węże, sfenodonty i Choristodera[62].
Przetrwałe linie ptaków Neornithes, wśród nich przodkowie dzisiejszych bezgrzebieniowych, Galloanserae i różnorodne siewkowe, różnicowały się gwałtownie na początku paleogenu, zajmując nisze ekologiczne opuszczone przez wymarłe grupy dinozaurów, jak nadrzewne Enantiornithes, wodne Hesperornithes, czy nawet większe teropody: powstały Gastornis (Diatryma), Dromornithidae i Phorusrhacidae, zwane po angielsku „terror birds”. Szybkiemu różnicowaniu uległy także ssaki, które wyparły Neornithes z większości nisz ekologicznych na lądzie[63].
Systematyka
edytuj
Dinozaury należą do archozaurów, jak współczesne krokodyle. W obrębie gadów naczelnych dinozaury wyróżnia najbardziej ich sposób chodzenia. Ich kończyny leżą prosto pod tułowiem, podczas gdy łapy krokodyli czy jaszczurek są rozstawione na boki.
Dinozaury stanowią klad, obejmujący według większości analiz filogenetycznych dwie główne gałęzie: dinozaury gadziomiedniczne i dinozaury ptasiomiedniczne. Gadziomiedniczne obejmują taksony, których ostatni wspólny przodek z ptakami żył później niż ostatni wspólny przodek z ptasiomiednicznymi. Ptasiomiednicze znowu mają ostatniego wspólnego przodka z triceratopsem późniejszego niż z gadziomiednicznymi. Anatomicznie te dwie grupy w sposób najłatwiej zauważalny różni budowa miednicy. Wczesne gadziomiedniczne, czyli Saurischia – od wywodzących się ze starożytnej greki słów sauros (σαῦρος) oznaczającego jaszczura i ischion (ἰσχίον) znaczącego biodro – zachowały strukturę stawu biodrowego ich przodków. Ich kość łonowa kieruje się dogłowowo (do przodu)[64]. Ta podstawowa forma została zmodyfikowana przez rotację miednicy w tył o różne kąty u kilku grup (herrerazaur[65], terizinozauroidy[66] dromeozaury[67] i ptaki[58]). Saurischia obejmują teropody, wyłącznie dwunożne, o szerokim wachlarzu diet, oraz zauropodomorfy, długoszyich roślinożerców, w tym zaawansowane grupy czworonożne.
Poprzez kontrast ptasiomiedniczne, których nazwa, stworzona przez Harry’ego Seeleya, wywodzi się od greckich słów ornitos znaczącego „ptasi” i ischion, mają miednicę powierzchownie przypominającą ptasią. Kość łonowa przynajmniej częściowo kieruje się doogonowo[68], mniej więcej równolegle do kości kulszowej (położenie opistopubiczne). Inną cechą wyróżniającą tę grupę jest kość przedzębowa[69].
Podział dinozaurów na dwie grupy siostrzane – ptasiomiedniczne i gadziomiedniczne – został podważony przez Barona, Normana i Barretta (2017) oraz Cau (2018); z przeprowadzonych przez autorów analiz filogenetycznych wynika bliższe pokrewieństwo teropodów z dinozaurami ptasiomiednicznymi niż z zauropodomorfami[28][70]. Langer i współpracownicy (2017) w wyniku przeprowadzenia analiz filogenetycznych opartych na zmodyfikowanej macierzy danych z analizy Barona, Normana i Barretta (2017) i uwzględniających większą liczbę taksonów uzyskali drzewa filogenetyczne potwierdzające tradycyjny podział dinozaurów na gadziomiedniczne i ptasiomiedniczne. Z analiz tych wynika jednak, że wystarczy wydłużenie drzewa filogenetycznego o dwa lub trzy stopnie w stosunku do najbardziej parsymonicznego, aby uzyskać drzewo, według którego teropody są bliżej spokrewnione z ptasiomiednicznymi niż z zauropodomorfami; z kolei aby uzyskać drzewo, według którego zauropodomorfy są bliżej spokrewnione z ptasiomiednicznymi niż z teropodami, konieczne jest wydłużenie drzewa filogenetycznego o cztery stopnie w stosunku do najbardziej parsymonicznego[71].
Dinozaury a ptaki – problem z zaklasyfikowaniem
edytujOd ponad wieku naukowcy zdawali sobie sprawę, że wiele dowodów wskazuje na bliskie pokrewieństwo ptaków i jednej z grup archozaurów – jedyną obecnie żyjącą grupą siostrzaną ptaków są należące do archozaurów krokodyle[72][73]. Gdyby ranga grupy systematycznej i pozycja w hierarchii systemu linneuszowskiego zależała tylko od pokrewieństwa, jak tego chcą kladystycy, to wówczas zgodnie ze spostrzeżeniami niektórych paleontologów ptaki należałoby umieścić wśród należących do archozaurów teropodów (tzn. wśród dinozaurów)[74]. Byłoby to jednak niedogodne z powodów praktycznych: ptaki to bogata w gatunki grupa czworonogów (Tetrapoda), jej przedstawiciele wykazują duże zróżnicowanie pod względem ekologicznym. Przypisanie ptakom rangi niższej od gromady, w nadrzędzie dinozaurów, ograniczyłoby możliwości odzwierciedlenia tego zróżnicowania w systematyce[72]. Jednak wśród paleontologów brak zgody co do tego, która grupa gadów jest najbliższa ptakom – niektórzy z nich kwestionują pochodzenie ptaków od teropodów, wskazując na brak jednoznacznych dowodów umożliwiających rozstrzygnięcie wzajemnych relacji filogenetycznych wśród archozaurów[1]. Ponadto ptaki, w związku z uzyskaniem zdolności do lotu i związaną z tym gruntowną przebudową ciała, wykazują wiele cech wspólnych, a ich budowa odróżnia je wyraźnie od pozostałych archozaurów[72]. Jednak najwcześniejsze ptaki i spokrewnione z nimi nieptasie teropody także wykazują wiele podobieństw, a niektóre cechy uznawane za wyróżniające ptaki (np. obecność piór) pojawiały się także u innych grup nieptasich dinozaurów[73][75]. Wielu zoologów z różnych powodów zdaje się nie zauważać pojawiających się coraz nowych dowodów na pochodzenie ptaków od archozaurów, czy też dokładniej – od teropodów[74]. Powoduje to problemy w wypracowaniu wspólnego stanowiska pomiędzy filogenetykami-kladystami i systematykami. Nieortodoksyjną propozycję konsensusu przedstawił Benton: przy zachowaniu rangi ptaków jako gromady, zasugerował on włączenie jej jako jednostki podrzędnej do nadrzędu Dinosauria (a więc niższej niż gromada rangi systematycznej)[76]. Konsekwencją braku powszechnie przyjętego stanowiska w sprawie umiejscowienia ptaków w hierarchii systematycznej jest to, że w podręcznikach, bazach informacji taksonomicznej i encyklopediach, ptaki cały czas są traktowane jako niezależna od dinozaurów gromada w obrębie czworonogów (Tetrapoda)[77][78][79].
-
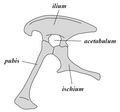 Miednica gadziomiednicznych widziana od lewej
Miednica gadziomiednicznych widziana od lewej -
 Jej przykład stanowi miednica tyranozaura
Jej przykład stanowi miednica tyranozaura -
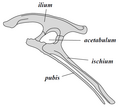 Miednica ptasiomiednicznych
Miednica ptasiomiednicznych -
 Np. edmontozaura
Np. edmontozaura




Taksonomia
edytujPoniżej podano uproszczoną klasyfikację głównych grup dinozaurów w oparciu o pokrewieństwo ewolucyjne na podstawie listy dinozaurów mezozoicznych autorstwa Holtza (2008)[2]. Krzyżykiem (†) oznaczono grupy wymarłe.
- Dinozaury – Dinosauria
- Dinozaury gadziomiedniczne – Saurischia
- Teropody –Theropoda (dwunożne, w większości mięsożerne)
- †Herrerazaury – Herrerasauria (wcześni dwunożni mięsożerny). Niekiedy wyróżnia się prócz Herreresauridae rodzinę staurikozaurów, wyłączając obie z teropodów i uznając za bazalne gadziomiedniczne[80]
- †Celofyzoidy – Coelophysoidea (niewielkie, dwunożne teropody, celofyz i jego bliscy krewni)
- †Dilofozaury – Dilophosauridae (wczesne mięsożerne teropody z grzebieniami na głowach)
- †Ceratozaury – Ceratosauria (dominujący południowi mięsożercy kredy z rogami na głowach)
- Tetanury – Tetanurae (najliczniejsza grupa teropodów)
- †Megalozauroidy – Megalosauroidea (wczesna grupa wielkich mięsożerców obejmująca spinozaury o częściowo wodnym trybie życia)
- †Karnozaury – Carnosauria (zwierzęta pokrewne allozaurowi i karcharodontozaurowi)
- Celurozaury – Coelurosauria (opierzone teropody o wielkiej różnorodności wielkości ciała i zajmowanej niszy)
- †Kompsognaty – Compsognathidae (pospolite wczesne celurozaury o zredukowanych przednich łapach)
- †Tyranozaury – Tyrannosauridae (tyranozaur i jego bliscy krewni o zredukowanych przednich łapach)
- †Ornitomimozaury – Ornithomimosauria (przypominający strusie w większości bezzębni mięsożercy bądź możliwi roślinożercy)
- †Alwarezaury – Alvarezsauroidea (niewielcy owadożercy o zredukowanych przednich kończynach noszących jeden wyrośnięty pazur)
- Maniraptory – Maniraptora („ręczni łowcy” o długich, smukłych przednich łapach i palcach)
- †Therizinosauria (dwunożni roślinożerny o wielkich pazurach na rękach i małych głowach)
- †Owiraptorozaury – Oviraptorosauria (w większości bezzębne, o niepewnym pożywieniu i stylu życia)
- †Archaeopterygidae (małe, uskrzydlone teropody, prymitywne ptaki)
- †Deinonychozaury – Deinonychosauria (małe bądź średnich rozmiarów przypominające ptaki, wydatny pazur na stopie)
- Avialae (współczesne ptaki i ich wymarli krewni)
- †Scansoriopterygidae (małe, prymitywne Avialae, długi trzeci palec)
- †Omnivoropterygidae (wczesne Avialae o krótkim ogonie)
- †Confuciusornithidae (niewielkie, bezzębne)
- †Enantiornithes (prymitywni, latający mieszkańcy drzew)
- Euornithes (zaawansowane ptaki latające)
- †Yanornithiformes (bezzębne chińskie ptaki z kredy)
- †Hesperornithes (wyspecjalizowane ptaki nurkujące w wodzie)
- Ptaki – Aves (współczesne ptaki posiadające dziób i ich wymarli krewni)

Zauropody z grupy Macronaria: kamarazaur, brachiozaur, Giraffatitan i euhelop
- †Zauropodomorfy – Sauropodomorpha (głównie roślinożercy o niewielkich głowach, długich szyjach, długich ogonach)
- †Guaibasauridae (formy małe, prymitywne, wszystkożerne)
- †Plateozaury – Plateosauridae (prymitywne prozauropody, dwunożne)
- †Riojasauridae (nieduże, prymitywne)
- †Masospondyle – Massospondylidae (nieduże, prymitywne)
- †Zauropody – Sauropoda (bardzo duże i ciężkie, zazwyczaj ponad 15 m długości, czworonożne)
- †Wulkanodony – Vulcanodontidae (prymitywne zauropody o słupowatych kończynach)
- †Eusauropoda („właściwe zauropody”)
- †Cetiozaury – Cetiosauridae („wielorybie jaszczury”)
- †Turiazaury – Turiasauria (europejska grupy jurajskich i kredowych zauropodów)
- †Neosauropoda („nowe zauropody”)
- †Diplodokokształtne – Diplodocoidea (czaszki i ogony wydłużone, zęby typowo wąskie, ołówkowate)
- †Macronaria (kanciasta czaszka, zęby łyżkowate bądź ołówkowate)
- †Brachiozaury – Brachiosauridae (długa szyja, długie kończy przednie)
- †Tytanozaury – Titanosauria (grupa zróżnicowana; przysadziste, o szerokich biodrach, najpospolitsze w kredzie późnej na kontynentach południowych)

Przedstawiciele ornitopodów i heterodontozaurów: od lewej kamptozaur, iguanodon, szantungozaur (z tyłu), driozaur (z przodu), heterodontozaur, tenontozaur
- Teropody –Theropoda (dwunożne, w większości mięsożerne)
- †Dinozaury ptasiomiedniczne – Ornithischia (grupa zróżnicowana, głównie roślinożercy dwu- bądź czworonożni)
- †Heterodontozaury – Heterodontosauridae (niewielkie, bazalne formy roślinożerne bądź wszystkożerne o wydatnych zębach przypominających kły)
- †Tyreofory – Thyreophora (uzbrojone dinozaury w większości czworonożne)
- †Ankylozaury – Ankylosauria (pancerz kostny z łusek, niekiedy buława kostna na ogonie)
- †Stegozaury – Stegosauria (kolce i płyty)
- †Neornithischia („nowe ptasiomiedniczne”)
- †Ornitopody – Ornithopoda (dwu- bądź czworonożne, wykształciły sposób rozdrabniania pokarmu przy użyciu ruchliwej czaszki i licznych zębów)
- †Marginocefale – Marginocephalia (rozrośnięta czaszka)
- †Pachycefalozaury – Pachycephalosauria (dwunożne, czaszka tworzy kopułę bądź rozrasta się guzowato)
- †Ceratopsy – Ceratopsia (głównie czworonożne, kryza kostna, często także rogi)
- Dinozaury gadziomiedniczne – Saurischia


Badania uwzględniające 277 rodzajów dinozaurów opublikowane w 2002 przez Pisaniego, Yatesa, Langera i Bentona zaowocowały przedstawieniem następujących powiązań[81] (zmodyfikowano):







.jpg)
Dinozaury ptasiomiedniczne (Ornitischia)
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||



Zauropodomorfy (Sauropodomorpha)
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||





.png)

Teropody (Theropoda)
Biologia
edytujWiedza o dinozaurach pochodzi z badań różnorodnych szczątków kopalnych i innych. Zaliczają się tutaj sfosylizowane kości, kał, tropy, gastrolity, pióra, odciski skóry, trzewia i tkanki miękkie[82][83]. Do zrozumienia dinozaurów przyczynia się wiele dziedzin nauki, jak fizyka, a w szczególności biomechanika, chemia, biologia, nauki o Ziemi, do których zalicza się też paleontologia.
Wielkość
edytuj
Obecnie potwierdzono zróżnicowanie średniej wielkości dinozaurów w triasie, jurze wczesnej, jurze późnej i kredzie[49]. Drapieżne teropody, zajmujące większość nisz lądowych mięsożerców w mezozoiku, najczęściej cechowały się masą w przedziale od 100 do 1000 kilogramów, jeśli posortować je wedle rzędu wielkości, podczas gdy ssaki drapieżne holocenu przypadają 1 rząd wielkości niżej (10-100 kilogramów)[84]. Modalna masy mezozoicznych dinozaurów leży pomiędzy 1 a 10 tonami[85]. Kontrastuje to ostro ze ssakami kenozoiku: Muzeum Historii Naturalnej w Waszyngtonie szacuje je na pomiędzy 2 i 5 kilogramów[86].
Największymi i najcięższymi z dinozaurów były zauropody. Przez większą część ery dinozaurów najmniejsze zauropody przerastały jakiekolwiek inne zwierzęta w ich środowiskach. Największe z nich z kolei były rzędu wielkości niespotykanego w przypadku żadnego innego zwierzęcia, jakie kiedykolwiek chodziło po ziemi. Wielkie ssaki prehistoryczne (megafauna), jak największy lądowy ssak w historii planety Paraceratherium, wydawały się karłami w porównaniu z olbrzymimi zauropodami. Jedynie żyjące i współcześnie wieloryby dorównują im, a nawet przewyższają je wielkością[87].
Wielkość a żywienie się
edytujZaproponowano kilka różnych korzyści odnoszonych dzięki gigantycznym rozmiarom zauropodów. Duże rozmiary wiążą się z mniejszym zagrożeniem ze strony drapieżników. Zmniejszają zużycie energii na jednostkę masy ciała. Wiążą się z długowiecznością[21]. Istnieją też pewne ogólne prawidłowości. Większe zwierzęta zamieszkują obszary o większej powierzchni. Roślinożercy są więksi od mięsożerców, bowiem pokarm roślinny dostępny jest w większej obfitości niż mięso. Zwierzęta zmiennocieplne, cechujące się wolniejszym metabolizmem i wymagające przez to mniej pożywienia, mogą osiągnąć większe rozmiary niż stałocieplne. O ile ta ostatnia kwestia w przypadku zauropodów jest dyskusyjna, to duże stężenie dwutlenku węgla w atmosferze w mezozoiku wiązało się z bujnym wzrostem roślin, zapewniającym obfite źródło pokarmu olbrzymim zauropodom[88]. Ken Carpenter dużą rolę przypisuje czynnikowi związanemu z żywieniem się[21].
Hummel i Clauss uznają zauropody za grupę dużych, lądowych roślinożerców, która odniosła największy sukces ewolucyjny. Jednak o sposobie ich żywienia wiadomo niewiele. Układ pokarmowy nie zachowuje się w zapisie kopalnym, gastrolity trudno przyporządkować zauropodom. W efekcie wiedza na ten temat pochodzi głównie z porównań ze współcześnie żyjącymi organizmami. Poza tym zauropody nie były jednorodną grupą, jeśli chodzi o żywienie się. Prawdopodobnie pochłaniały duże ilości pokarmu. W przypadku osobników dorosłych był to wyłącznie pokarm roślinny (skrzypowe, paprotniki, nagonasienne), o czym świadczy budowa zębów, ołówkowatego kształtu u diplodoków i tytanozaurów, a bardziej masywnych, łyżkowatych u bazalnych zauropodów i bazalnych Macronaria. Dzięki długim szyjom zwierzęta mogły żywić się pokarmem roślinnym dostępnym na znacznej wysokości. Roślinożercy wykazują tendencję do żywienia się pokarmem, którego większą ilość mogą zjeść dzięki mniejszej liczbie kęsów. Drzewa iglaste wydają się tu lepszy pokarmem niż liściaste czy paprotniki. Osobniki, które dopiero co się wylęgły, mogły okazjonalnie żywić się stawonogami bądź innymi drobnymi zwierzętami. Pokarm nie był żuty, podobnie jak u stegozaurów i dzisiejszych ptaków. Być może ulegał wstępnemu rozdrobnieniu w żołądku, jak u ptaków, jednak nie ma to zgody wśród badaczy. Bardzo długo zalegał w układzie pokarmowym. W ich jelitach prawdopodobnie bytowały bakterie symbiotyczne, uczestniczące w trawieniu pokarmu, a być może również grzyby[89]. Carpenter uważa, że duże zwierzęta wydajniej trawią niż małe, właśnie dlatego, że pokarm spędza więcej czasu w ich długich przewodach pokarmowych. Pozwala im to w jego opinii przetrwać dzięki żywności o mniejszej wartości odżywczej niż w przypadku stworzeń niewielkich. Pozostałości zauropodów znajdywano głównie w formacjach górskich, odpowiadających terenom suchym lub przynajmniej okresowo suchym[21]. W zapisie kopalnym towarzyszą im iglaste (tak jak ornitopodom okrytonasienne)[89]. Zdolność pochłaniania olbrzymich ilości ubogiego w składniki odżywcze pożywienia była w takich środowiskach korzystna[21]. Z drugiej strony konieczność dostarczenia organizmowi dużej ilości pożywienia ogranicza rozmiary zwierzęcia, zwłaszcza zauropoda o niewielkiej głowie, bowiem wraz z rozmiarami wzrasta czas, który zwierzę musi poświęcić na jedzenie[89].
Największy i najmniejszy
edytujNauka prawdopodobnie nigdy nie zdobędzie pewności, który z żyjących kiedykolwiek dinozaurów był największy, a który najmniejszy. Tylko niewielki ułamek prehistorycznych zwierząt zachował się dzięki fosylizacji, a większość z pozostałości spoczywa zagrzebana w ziemi. Bardzo nieliczne okazy zostają wydobyte jako kompletne szkielety, a odciski skóry czy pozostałości innych tkanek miękkich są rzadkie. Odtwarzanie kompletnych szkieletów poprzez porównywanie wielkości i budowy kości z należącymi do podobnych, acz lepiej poznanych gatunków jest nieścisłe, a rekonstrukcja mięśni i innych narządów żyjących zwierząt to w najlepszym razie spekulacje.

Najwyższym i najcięższym spośród dobrze zachowanych szkieletów należy do Giraffatitan brancai (dawniej klasyfikowanego jako gatunek brachiozaura). Jego szczątki odkryto w Tanzanii pomiędzy 1907 i 1912. Kości kilku osobników podobnej wielkości połączono w pojedynczy szkielet, zmontowany i wystawiany w Muzeum Historii Naturalnej w Berlinie[90]. Jego wysokość wynosi 12 m, podczas gdy długość – 22,5 m. Masę zwierzęcia szacuje się na 30-60 ton (dokładniejsze oszacowanie podaje 39,5 tony[91]).
Istniały jeszcze większe dinozaury, ale wiedza o nich bazuje całkowicie na nielicznych lub fragmentarycznych skamielinach. Większość z największych okazów roślinożerców odkryto w latach siedemdziesiątych XX wieku lub później. Zalicza się doń masywny argentynozaur, który mógł ważyć od 80 do 100 ton[21]. Wedle innych danych ważył on jednak mniej, 73 tony[88]. Jednak nawet przez przyjmujących to ostrożniejsze oszacowanie uznawany bywa za największego z dinozaurów. Drugie miejsce zajmuje Antarctosaurus giganteus, 69-tonowy olbrzym również zamieszkujący Amerykę Południową[91]. Do najdłuższych należą osiągający 33,5 m przedstawiciele Diplodocus hallorum[21], wcześniej określani mianem Seismosaurus, a także mierzący 33 m długości superzaur[92] i najwyższy, osiemnastometrowy zauroposejdon, który sięgnął by szóstego piętra. Najcięższym i najdłuższym mógł być jednak amficelias (Amphicoelias fragillimus), znany jedynie z utraconego niekompletnego łuku kręgowego opisanego w 1878. Ekstrapolując na podstawie ilustracji tego fragmentu kości, długość zwierzęcia szacuje się 58 m, jego masę zaś na 120-260 ton[21]. Wspomniany zauroposejdon ważył 55 ton, afrykański Paralititan 59 ton[88]. Największym znanym teropodem był z kolei spinozaur. Osiągał długość 16–18 m oraz masę 8150 kg[93]. Wielkimi mięsożercami byli także giganotozaur, karcharodontozaur i tyranozaur[94]. Ważyły one odpowiednio 9, 7,5 oraz 6,25 tony[88].
Jeśli nie brać pod uwagę ptaków, najmniejsze znane dinozaury były wielkości gołębia[95]. Najmniejsze nieptasie dinozaury należały do teropodów blisko spokrewnionych z ptakami. Na przykład długość szkieletu anchiornisa (Anchiornis huxleyi) wynosiła niecałe 35 cm[95][96]. A. huxleyi jest obecnie najmniejszym znanym nieptasim dinozaurem opisanym na podstawie szkieletu osobnika dorosłego. Jego masę szacuje się na 110 gramów[96]. Najmniejsze roślinożerne nieptasie dinozaury to mikroceratus i wannanozaur. Oba mierzyły około 60 cm długości[2][97] i zaliczały się do marginocefali.
Zachowanie
edytuj
Wiele współczesnych ptaków prezentuje zachowania wysoce społeczne, często żyją w stadach. Panuje ogólna zgoda, że niektóre zachowania częste u ptaków, jak też u krokodyli (najbliższych dzisiejszych krewnych ptaków) były też częste u wymarłych grup dinozaurów. Interpretacje zachowania gatunków kopalnych bazują generalnie na pozach szkieletów, ich siedliskach, symulacjach komputerowych biomechaniki i porównaniach ze współczesnymi zwierzętami zajmującymi podobne nisze ekologiczne.
Pierwsze potencjalne potwierdzenie szeroko rozpowszechnionych zachowań stadnych wśród wielu grup nieptasich dinozaurów zdobyto w 1878, gdy odkryto 31 Iguanodon bernissartensis. Sądzi się, że te ornitopody zginęły razem w miejscu dzisiejszego Bernissart na terenie Belgii, po tym, jak wpadły w głęboki, zalany wodą lej krasowy, gdzie utonęły[98]. Odkrywano później kolejne miejsca masowych śmierci. Znalezisko z Sauerlandu, w którym prócz przeważających I. atherfieldensis spotyka się także I. bernissartensis, obejmujące także młode osobniki, interpretuje się jako osobniki zebrane w jednym miejscu przez powódź bądź jako stado przekraczające rzekę[99]. Wraz z licznymi tropami sugeruje to, że zachowania stadne były częste u wielu wczesnych gatunków dinozaurów. Ślady setek, a może i nawet tysięcy roślinożerców wskazują, że hadrozaury mogły tworzyć wielkie stada, jak dziś bizony w Ameryce czy afrykańskie springboki. Tropy zauropodów dokumentują przemieszczanie się tych zwierząt w grupach składających się z osobników kilku różnych gatunków. Wymienić można tutaj znalezisko z Oxfordshire w Anglii[100]. Nie potwierdza to jednak tworzenia właściwych struktur stadnych[101]. Gromadzenie się w stada mogło wyewoluować z uwagi na ochronę członków, w tym młodych, czy też migracje. Potwierdzono, że niedojrzałe osobniki z różnych grup wolno rosnących dinozaurów, w tym różne teropody, zauropody, ankylozaury, ornitopody i ceratopsy, tworzyły zgrupowania. Przykład stanowi miejsce w Mongolii Wewnętrznej, gdzie spoczywają szczątki ponad 20 sinornitomimów. Pozostałości spoczywają w jednej płaszczyźnie, na powietrzu leżały poniżej jednego roku. Zwierzęta zmarły w takich samych warunkach, a ich zwłoki szybko zostały przykryte. Zwierzęta w momencie wspólnego zgonu liczyły od roku do lat siedmiu, były to więc osobniki młodociane lub podrostki. Nie było natomiast piskląt ani całkiem dorosłych zwierząt. Zgromadzenie to interpretuje się jako grupę społeczną uwięzioną w błocie. Znajdywano również grupy osobników młodocianych falkariusów, pinakozaurów, diplodoków, ornitopodów i ceratopsów. Przykłady masowej śmierci współczesnych zwierząt generalnie wiążą się z życiem stadnym, jednakże taki tryb życia u dzisiejszych archozaurów jest rzadkością. Spotyka się go u ptaków późno osiągających dojrzałość płciową, jak strusie czy nandu szare, kruki zaś wspólnie żerują i wykorzystują wspólne grzędy. Zwraca się też uwagę, że sinornitomim był prawdopodobnie roślinożerny i w związku z tym cechował się w młodym wieku znaczną umieralnością[102]. Interpretacja dinozaurów jako stworzeń stadnych rozszerzyła się na mięsożerne teropody przedstawiane jako grupowych łowców, współpracujących w celu pokonania większej zdobyczy[103][104]. Jednakże taki sposób życia jest rzadki wśród współczesnych ptaków, krokodyli i innych kręgowców określanych mianem gadów, a przesłanki tafonomiczne sugerujące polowania w grupie na podobieństwo ssaków w przypadku takich teropodów, jak deinonych czy allozaur można również interpretować jako rezultat śmiertelnych utarczek pomiędzy pożywiającymi się zwierzętami, jakie zdarzają się wśród współczesnych polujących diapsydów[105].

Grzebienie i kryzy pewnych dinozaurów, jak marginocefale, teropody i lambeozauryny, mogły być zbyt delikatne, by służyć do obrony. W związku z tym pełniły raczej rolę dekoracyjną w pokazach seksualnych czy groźbach. Jednakże mało wiadomo na temat łączenia się w pary czy terytorializmu nieptasich dinozaurów. Rany głowy powstałe na skutek ugryzień wskazują, że przynajmniej teropody angażowały się w aktywne konfrontacje bitewne[106].
Z behawioralnego punktu widzenia jedną z najbardziej wartościowych skamieniałości znaleziono na pustyni Gobi w 1971. Obejmowała ona welociraptora atakującego protoceratopsa[107]. Dowodzi ona, że dinozaury naprawdę się atakowały[108]. Dodatkowo atakowanie żywej zdobyczy potwierdza częściowo wygojony ogon kaczodziobego edmontozaura. Jego uszkodzenie wskazuje, że został on ugryziony przez tyranozaura, ale przeżył[108]. Potwierdzono również kanibalizm wśród niektórych gatunków dinozaurów. Ślady zębów znalezione na Madagaskarze w 2003 potwierdzają go u teropoda mażungazaura[109].
Porównania pierścieni twardówkowych nieptasich dinozaurów, ptaków nowoczesnych i zwierząt określanych mianem gadów ujawniło dzienną aktywność dinozaurów. Choć zasugerowano, że większość dinozaurów było aktywnych za dnia, porównania wykazały, że małe drapieżne dinozaury, jak dromeozaury, jurawenator i megapnozaur, prowadziły nocny tryb życia. Dużych i średnich rozmiarów roślinożercy i wszystkożercy, jak ceratopsy, zauropodomorfy, hadrozaury, ornitomimozaury, mogły być aktywne podczas krótkich okresów za dnia. Mały ornitopod agilizaur prowadził dzienny tryb życia[110].
Bazując na niedawnych dowodach kopalnych (oryktodrom), uważa się pewne ornitopody za grzebiące w ziemi[59]. Wiele ptaków nowoczesnych wiedzie nadrzewny tryb życia. Podobnie czyniły ptaki mezozoiku, zwłaszcza Enantiornithes[111]. Pewne wczesne zbliżone do ptaków gatunki także mogły już prowadzić życie nadrzewne, jak dromeozaury, np. mikroraptor[112]). Jednak większość nieptasich dinozaurów zdawała się chodzić po gruncie. Dobre zrozumienie przemieszczania się dinozaurów stanowi klucz do tworzenia modeli ich zachowania. Biomechanika w szczególności pozwoliła wniknąć w to zagadnienie. Dla przykładu badania sił rozwijanych przez mięśnie i ciężaru ich szkieletów pozwalały na oszacowanie prędkości biegu[113], możliwości tworzenia przez diplodoki gromu dźwiękowego dzięki biczowatemu ogonowi[114], a także możliwości pływania zauropodów[115].
Komunikacja
edytuj
Ptaki nowoczesne znane są z komunikowania się ze sobą – stosują głównie sygnały wizualne i dźwiękowe. Duża różnorodność struktur służących pokazom u kopalnych grup dinozaurów wskazuje, że komunikacja wzrokowa zawsze odgrywała wśród dinozaurów ważną rolę. Jednak ewolucja wokalizacji wśród dinozaurów jest mniej pewna. W 2008 paleontolog Phil Senter zbadał dowody na wokalizację u zwierząt z mezozoiku, w tym dinozaurów[116]. Doszedł do wniosku stojącego w sprzeczności z popularnym w animacjach wizerunkiem ryczących dinozaurów – prawdopodobnie większość mezozoicznych dinozaurów nie potrafiła wydawać z siebie żadnych dźwięków. Jednak lambeozaury dzięki swym wydrążonym grzebieniem pełniącym funkcje komór rezonansowych wydawały z siebie szeroką gamę odgłosów[117][118]. By dojść do takiej konkluzji, Senter przestudiował rozmieszczenie narządów głosowych u współczesnych gadów i ptaków. Zaobserwował, że struny głosowe w krtani wyewoluowały prawdopodobnie wielokrotnie wśród zwierząt określanych gadami. Wymienić można krokodyle, zdolne do wydawania gardłowych dźwięków. Z drugiej strony, u ptaków odgłosy powstają w specjalnym narządzie, występującym tylko u nich, zwanym krtanią tylną (syrinx), niespokrewnionym z krtanią. Jest to chrzęstna struktura leżąca w miejscu połączenia tchawicy i oskrzeli głównych, zaopatrzona we wpuklające się do światła fałdy, których drgania powodują powstawanie dźwięków. Narząd ten wyewoluował niezależnie z narządów głosu ptasich przodków. Działanie syrinx zależy od systemu worków powietrznych. W szczególności wymaga on istnienia worka obojczykowego niedaleko widełek bądź obojczyka. Występowanie tego worka wiąże się z istnieniem obszernych otworów w kościach, w tym wyróżniającego się otworu w kości ramiennej. Obszerny układ worków powietrznych pozostaje unikalną cechą dinozaurów gadziomiednicznych, jednak worek obojczykowy konieczny do wydawania dźwięków stanowi cechę późną, powstałą między innymi przez połączenie dwóch innych worków powietrznych. Nie występuje on w zapisie kopalnym przed pojawieniem się Enantiornithes. Jedynym znanym wyjątkiem jest Aerosteon o spneumatyzowanych widełkach obojczyka, który prawdopodobnie wykształcił worek obojczykowy niezależnie od ptaków z przyczyn innych niż wokalizacja[116].

Najprymitywniejszymi dinozaurami, u których potwierdzono istnienie wydającej dźwięki syrinx, są Enantiornithes, zaliczane już do ptaków. Żadne archozaury z ptasiej linii bardziej prymitywne od wymienionych nie wydawały z siebie odgłosów. Raczej różnego rodzaju argumenty wskazują na używanie przez wczesne dinozaury głównie komunikacji wzrokowej w formie wyróżniających się wyglądem i prawdopodobnie żywo zabarwionych rogów, kryz, grzebieni, żagli i piór. Przypominają w tym względzie niektóre współczesne grupy gadów, jak jaszczurki, u których wiele form jest w znacznym stopniu cichych (choć jak dinozaury dysponują one dobrze rozwiniętym zmysłem słuchu), ale wykorzystujących złożone ubarwienie i wystawiających się na pokaz dla komunikacji[116]. Nieptasie dinozaury wykorzystywały inne metody tworzenia dźwięków dla komunikowania się. Inne zwierzęta, w tym inne gady, używają różnorodnych dźwięków niewokalnych: syczą, kłapią bądź zgrzytają jamą ustną, wykorzystują swe otoczenie, np. pluskają i uderzają skrzydłami (mogły tak czynić uskrzydlone maniraptory)[116].
Rozmnażanie
edytuj
Wszystkie dinozaury składają jaja zawierające błony płodowe. Do 2020 roku znane były tylko jaja dinozaurów otoczone twardą osłonką zbudowaną głównie z węglanu wapnia[119]. Badania Norella i współpracowników (2020) nad jajami protoceratopsów i muszaurów sugerują jednak, że jaja tych dinozaurów miały miękką osłonę; zdaniem powołanych autorów jaja pierwszych dinozaurów były pozbawione twardej wapiennej osłonki, która wykształciła się niezależnie u co najmniej trzech grup dinozaurów[120]. Jaja składane są zwykle w gnieździe. Większość gatunków buduje dość zawiłe gniazda, tworząc kopce, kopuły, płyty, zagłębienia, jamy[121]. Pewne gatunki ptaków nowoczesnych nie tworzą gniazd. Zamieszkujący klify nurzyk zwyczajny składa jaja na nagiej skale, a samiec pingwina cesarskiego trzyma jaja pomiędzy swymi stopami a tułowiem. Prymitywne ptaki i wiele nieptasich dinozaurów często składało jaja we wspólnych gniazdach. Inkubacją jaj opiekowały się głównie samce. O ile samice ptaków nowoczesnych dysponują tylko jednym funkcjonalnym jajowodem, bardziej prymitywne ptaki i inne dinozaury miały dwa, jak krokodyle. Niektóre nieptasie dinozaury, jak troodon, składały jaja wielokrotnie: matka składała parę jaj co 1–2 dni. Jednoczesne wykluwanie się potomstwa zapewniano poprzez opóźnienie wysiadywania aż do chwili złożenia wszystkich jaj[122].
Publikacja Lee i Werning z 2008 dostarczyła informacji nie tylko na temat rozrodu, ale także dojrzewania dinozaurów. Bazuje ona na wcześniejszym stwierdzeniu pewnego szczególnego rodzaju tkanki kostnej, kości szpikowej (medullary bone), u tyranozaura. Autorzy pracy wykryli ją również u allozaura i tenontozaura. Tkanka tego rodzaju nie występuje w organizmie przez cały czas. U współczesnych ptaków (nie u wszystkich) pojawia się w jamach szpikowych przed owulacją, stanowiąc źródło wapnia dla tworzących się skorupek, po czy szybko zanika. Ponieważ nie występuje ona u krokodyli, uważa się, że rozwinęła się u Ornithodira w triasie późnym[123].
Pojawienie się tego rodzaju tkanki kostnej wiąże się ze spowolnieniem wzrostu osobniczego już po okresie największego wzrostu. Zbadana samica tenontozaura miała 8 lat, allozaura – 10 lat, tyranozaura zaś – 18 lat. Jednakże wobec tego kość szpikowa musiała pojawić się, zanim zwierzę osiągnęło swój maksymalny wzrost. Z badania tego wynika również, że nieptasie dinozaury różniły się tempem wzrostu od współczesnych gadów. Rosły szybciej, wcześnie przystępowały do rozrodu. Ogólnie zwierzęta większych rozmiarów rosną szybciej niż ich mniejsi krewni. Jednakże gdyby, z zachowaniem skali, nieptasie dinozaury rosły w tempie współczesnych gadów, tkanka taka powinna pojawić się lata, jeśli nie dekady później. Autorzy widzą więc w dinozaurach szybko rosnące zagniazdowniki, przystępujące do rozrodu po osiągnięciu połowy, a nawet tylko trzeciej części maksymalnych rozmiarów ciała. Umieralność osobników dorosłych oceniają bowiem jako wysoką – szybsze przystąpienie do rozrodu wiązało się z większą szansą na pozostawienie po sobie potomstwa. Przykładowo osiągnięcie wielkości dorosłego osobnika zajmowało najcięższym zauropodom kilka dekad, ale rozmnażać się mogły już w wieku około 19 lat. Strategia taka różni z kolei nieptasie dinozaury od dzisiejszych ptaków[123].

Wśród ptaków nowoczesnych rozpowszechniona jest również opieka rodzicielska nad młodymi po wykluciu. Jack Horner odkrył w 1978 gniazdowisko majazaury (której nazwa oznacza „dobrą jaszczurzą matkę”) w Montanie. Widać było ślady opieki rodzicielskiej kontynuowanej długo po wykluciu się młodych ornitopodów. Sugeruje to, że zachowanie takie mogło być częste wśród wszystkich, także nieptasich dinozaurów[124]. Istnieją dowody, że inne, niezaliczane do teropodów dinozaury, jak patagońskie tytanozaury, także gniazdowały w dużych grupach. Wskazuje na to odkrycie z 1997[125]. Mongolski owiraptor Citipati osmolskae odkryty został w 1993 w pozycji wysiadującego kurczaka. Wskazuje to, że wykorzystywał warstwę izolacyjną tworzoną przez pióra, by utrzymać jaja w cieple[126]. Częste występowanie opieki rodzicielskiej u wszystkich dinozaurów wsparły inne znaleziska. Na przykład skamieniałe szczątki grupy psitakozaurów zawierają pozostałości osobnika dorosłego i 334 młodocianych. W tym przypadku dużą liczbę młodych wyjaśnić może wspólne gniazdowanie[127]. Co więcej, znaleziono bezzębne zarodki dinozaurów należące do prozauropoda masospondyla. Oznacza to, że wymagały one pomocy dorosłego w dostarczaniu im pokarmu po wykluciu[128]. Ślady także potwierdzają zachowania rodzicielskie: choćby tropy ornitopodów na Isle of Skye w północno-zachodniej Szkocji[129]. Znaleziono gniazda i jaja wszystkich większych grup dinozaurów. Wydaje się prawdopodobne, że wszystkie dinozaury opiekowały się swym potomstwem przez pewien czas przed i niedługo po wykluciu.
Wydalanie
edytujJak inne gady, dinozaury są pierwotnie urykoteliczne. Znaczy to, że ich nerki filtrują i wydalają z krwiobiegu azotowe produkty przemiany materii w postaci kwasu moczowego, który przez moczowody dostaje się do jelita. U większości żyjących dziś gatunków kwas moczowy opuszcza organizm wraz z kałem w postaci półstałej[130][131][132]. Jednak przynajmniej niektóre ptaki nowoczesne, jak kolibry, są fakultatywnie amonioteliczne. Termin ten oznacza wydalanie większości azotowych produktów przemiany materii w postaci amoniaku[133]. Wydalają również kreatynę, bardziej niż kreatyninę, jak czynią ssaki. Razem z treścią jelit materiał ten wydostaje się z organizmu przez kloakę (stek)[134][135]. W dodatku wiele gatunków regurgituje wypluwki. Skamieniałe wypluwki, mogące pochodzić od dinozaurów, znane są od okresu kredowego[136].
Fizjologia
edytujPonieważ zarówno ptaki nowoczesne i krokodyle mają serce czterokomorowe, aczkolwiek zmodyfikowane u krokodyli, jest prawdopodobne, że cecha ta łączyła wszystkie archozaury, w tym wszystkie dinozaury[137].
Po odkryciu nieptasich dinozaurów paleontolodzy na początku uznali je za zmiennocieplne. Podejrzewana zimnokrwistość służyła do argumentowania za tym, że te dawne dinozaury były względnie wolnymi, niemrawymi stworzeniami, mimo że wiele współczesnych gadów porusza się szybko i zwinnie pomimo polegania na zewnętrznych źródłach energii cieplnej w regulacji temperatury swego ciała. Pomysł zmiennocieplnych, ociężałych dinozaurów stał się głównym punktem widzenia, nim Robert Bakker, wczesny zwolennik stałocieplności dinozaurów, opublikował swą wpływową publikację w 1968[138].
Obecnie dowiedziono bujnego rozwoju nieptasich dinozaurów i ptaków w klimacie o niskich temperaturach. W związku z tym przynajmniej niektóre z wczesnych gatunków musiały wewnętrznie regulować temperatury swych ciał. Większe gatunki wykorzystywały do tego swe rozmiary. Mniejszym pomagały okrywające ich ciała pióra. Dowody stałocieplności dinozaurów mezozoiku obejmują odkrycie polarnych dinozaurów australijskich i antarktycznych. Musiały one jakoś przetrwać ciemną, zimną zimę trwającą 6 miesięcy. Zawierają też analizę budowy naczyń krwionośnych skamieniałych kości, typowej dla zwierząt endotermicznych. Debata naukowa trwa, omawiając z kolei konkretne drogi, które zaprowadziły dinozaury ku wewnętrznej regulacji temperatur[139].
Układ oddechowy
edytuj
Omówiona powyżej kwestia metabolizmu wiąże się ściśle z budową układu oddechowego. Jednakże budujące go tkanki nie zachowują się w materiale kopalnym. Tymczasem u współcześnie żyjących ptaków obserwuje się charakterystyczną budowę płuc, zaopatrzonych w worki powietrzne. Jednak worki takie rzadko wnikają do kości. Pomimo tego udało się je znaleźć u allozauroida, którego Paul Sereno i współpracownicy nazwali Aerosteon. U gadziomiednicznych stwierdzono już wcześniej istnienie worków szyjnych, które jednak nie biorą udział w oddychaniu, u Aerosteon jednak stwierdzono istnienie worków komunikujących się z płucami, leżących zarówno wewnątrz klatki piersiowej, jak i poza nią (nawet w kości biodrowej). Odkrycie, że niektóre nieptasie teropody miały worki powietrzne połączone z płucami wskazuje, że ich płuca przypominały płuca współczesnych ptaków – sztywne i przytwierdzone do grzbietowej ściany klatki piersiowej[140].
Autorzy ci opisali również przebieg ewolucji worków powietrznych. W pierwszej fazie, najpóźniej na początku triasu późnego, pojawiły się worki szyjne, które następnie, w okresie jury, zróżnicowały się. Powstały worki związane z układem oddechowym. Przed końcem jury u maniraptorów wyewoluowała prymitywna pompa mostkowo-żebrowa, zaś pod koniec tego okresu można już mówić o zaawansowanej pompie. Powstanie worków wiązać się mogło nie tylko ze sprawniejszym oddychaniem, ale także z równowagą lokomotoryczną i termoregulacją[140].
Wczesne ptasie układy oddechowe były zdolne utrzymać wyższe tempo metabolizmu niż w przypadku ssaków podobnej wielkości i budowa taka utrzymała się. Poza zapewnianiem wydajnego dopływu tlenu szybki przepływ powietrza stanowił efektywny sposób chłodzenia ciała, kluczowy dla stworzeń aktywnych, ale zbyt dużych, by pozbyć się całego nadmiaru ciepła przez skórę[141].
Skóra
edytujW 2018 r. McNamara i inni wykazali, że dinozaury zrzucały drobne fragmenty skóry[142].
Powstanie ptaków
edytujMożliwość, że dinozaury są przodkami ptaków, zasugerował po raz pierwszy w 1868 Thomas Henry Huxley[b][143]. Po pracy Gerharda Heilmanna z początku XX wieku teorię dinozaurzego pochodzenia ptaków porzucono. Poparto pomysł, zgodnie z którym ptaki pochodzą od tekodontów. Kluczowym szczegółem potwierdzającym taką tezę był podejrzewany brak obojczyka u dinozaurów[144]. Jednakże późniejsze odkrycia pokazały, że obojczyki nie były wcale nieobecne w tej grupie, mogły one tworzyć pojedyncze złączone widełki[58]. Obojczyki dinozaurów odkryto już w 1924 u owiraptora, ale niesłusznie uznano je za interclavicle[145]. W latach siedemdziesiątych XX wieku John Ostrom powrócił do teorii dinozaurzego pochodzenia ptaków[146]. W następnych dekadach pogląd ten nabrał rozpędu w związku z nadejściem analizy kladystycznej[147] i wielkim postępem w odkryciach niewielkich teropodów i wczesnych ptaków[37]. Wspomnieć należy szczególnie skamieliny z formacji Yixian, gdzie znaleziono różnorodne nieptasie teropody i wczesne ptaki, w tym formy pokryte pewnego typu piórami[58]. Ptaki dzielą ponad setkę różnych cech anatomicznych z nieptasimi teropodami, które uznano ogólnie za ich najbliższych wymarłych krewnych[148].
Najbliższą ptakom grupę teropodów stanowią celurozaury maniraptory[58]. Mniejszość specjalistów, z których najbardziej znaczący są Alan Feduccia i Larry Martin, proponują inne ścieżki ewolucji ptaków, włączając w to zrewidowane wersje propozycji Heilmanna mówiącej o bazalnych archozaurach[149] lub tezę, jakoby maniraptory rzeczywiście były przodkami ptaków, ale same nie należały do dinozaurów (ich podobieństwo miałaby tłumaczyć konwergencja)[1].
Pióra
edytuj
Archaeopteryx to pierwsza skamieniałość, która ujawniła związki pomiędzy ptakami a dinozaurami (w starym znaczeniu). Uznaje się go za takson pośredni, formę przejściową wykazującą cechy obu grup. Został wydobyty na światło dzienne zaledwie 2 lata po ważnej publikacji Karola Darwina O powstawaniu gatunków. Jego odkrycie pobudziło rodzącą się dyskusję pomiędzy zwolennikami ewolucjonizmu a kreacjonizmu. Ten wczesny ptak tak bardzo przypomina nieptasie dinozaury, że przynajmniej jednego osobnika, w przypadku którego nie zachowały się odciski piór, mylnie uznano za kompsognata[150] (rodzina kompsognatów należy do celurozaurów daleko spokrewnionych z ptakami, dalej niż Arctometatarsalia, terizinozaury czy owiraptorozaury[151]).
Od lat dziewięćdziesiątych odkryto dużą liczbę kolejnych pierzastych dinozaurów. Zapewniło to silniejsze wsparcie tezy o bliskich pokrewieństwie pomiędzy dinozaurami w tradycyjnym rozumieniu a ptakami nowoczesnymi. Większość z takich osobników pochodzi z formacji Yixian z Liaoning w północno-wschodnich Chinach. W okresie kredy ziemie te stanowiły część kontynentu wyspowego. Choć pióra znaleziono tylko w kilku miejscach, możliwe, że nieptasie dinozaury żyjące gdziekolwiek na świecie także były upierzone. Brak szerokiego rozprzestrzenienia dowodów na taki stan rzeczy może wynikać z delikatnej budowy takich struktur, jak skóra czy pióra, które nieczęsto ulegają zachowaniu poprzez fosylizację, dlatego też brakuje ich w zapisie kopalnym. Do chwili obecnej protopióra, cienkie, nitkowate struktury, stwierdzono u bazalnych celurozurów, jak kompsognaty np. sinozauropteryks czy tyranozauroidy np. Dilong[152], ale haczykowate pióra znane są jedynie wśród maniraptorów, do których zaliczają się owiraptorozaury, troodonty, dromeozaury i ptaki[58][153]. Idea opierzonych dinozaurów nie obyła się bez kontrowersji. Być może najgłośniejsza krytyka popłynęła z ust Alana Feduccii i Theagartena Linghama-Soliara. Zaproponowali oni, że protopióra stanowią efekt rozkładu podskórnych włókien kolagenowych[154][155][156], a maniraptory o piórach haczykowatych nie były właściwie dinozaurami, reprezentując konwergentną z nimi linię ewolucyjną[1][155]. Jednakże ich punkt widzenia w znacznej części nie zyskał akceptacji innych badaczy do tego stopnia, że pojawiły się pytania o naukowość propozycji Feduccii w ogóle[157].
Tkanki miękkie
edytuj
Wielkie dinozaury mięsożerne miały skomplikowany system worków powietrznych, przypominający spotykany u ptaków nowoczesnych, zgodnie z badaniami przeprowadzonymi przez Patricka O’Connora z Uniwersytetu Ohio. Płuca teropodów, mięsożerców poruszających się na dwóch ptasiostopych nogach, prawdopodobnie łączyły się z workami powietrznymi w obrębie szkieletu, jak to ma miejsce u ptaków. O’Connor stwierdził, że to, co uznawano za unikalne dla ptaków, występowało również u niektórych ich przodków[158]. W publikacji z 2008 z pisma PLoS ONE opisano gatunek Aerosteon riocoloradensis. Jego szkielet zapewnia najsilniejsze dotychczas dowody na istnienie nieptasich dinozaurów o ptasim układzie oddechowym. Tomografia komputerowa skamielin Aerosteon ujawniły dowody na istnienie u niego worków powietrznych[140][159].
Argumenty behawioralne
edytujSkamieniałości troodontów Mei i sinornitoida pokazują, że niektóre nieptasie dinozaury spały z głową schowaną w ramionach[160]. Zachowanie to mogło pomagać w utrzymaniu ciepła. Jest ono charakterystyczne także dla ptaków nowoczesnych. Pewne okazy deinonychozaurów i owiraptorozaurów znaleziono zachowane na szczycie swych gniazd, prawdopodobnie wysiadujące jaja jak ptaki[161]. Współczynnik pomiędzy objętością jaja i masą ciała osobnika dorosłego tych dinozaurów sugeruje, że jaja były pierwotnie wysiadywane przez samca. Młode wydają się wysoko rozwinięte, podobnie jak u wielu ptaków nowoczesnych[162].
Niektóre nieptasie dinozaury używały kamieni jako żaren, jak to czynią ptaki nowoczesne. Zwierzęta połykają kamienie, wspomagając przez to trawienie i rozdrabnianie pokarmu w żołądku, w tym także twardych włókien roślinnych. Kamienie takie, określane mianem gastrolitów, znaleziono w połączeniu ze skamielinami[163].
Wymarcie większości grup
edytujOdkrycie, że ptaki są dinozaurami, doprowadziło do konstatacji, że dinozaury wcale nie wymarły, jak się powszechnie i błędnie uważa[164]. Jednakże wszystkie poza ptakami grupy dinozaurów, jak też wiele grup ptaków, rzeczywiście wyginęło dość nagle około 66 milionów lat temu razem z wieloma innymi grupami zwierząt. Wymienia się wśród nich amonity, mozazaury, plezjozaury, pterozaury i wiele grup ssaków[20]. Natomiast nie zauważono spadku populacji owadów, które mogły więc być dostępne jako źródło pokarmu. To masowe wymieranie określa się wymieraniem kredowym. Jego naturę badano obszernie od lat 70. XX wieku. Obecnie istnieje kilka powiązanych ze sobą teorii prezentujących jego wyjaśnienie. Panuje konsensus co do zderzenia Ziemi z innym ciałem niebieskim jako pierwotnej przyczyny wyginięcia nieptasich dinozaurów, pewni naukowcy podają też inne możliwe przyczyny bądź wspierają pogląd o nałożeniu się na siebie licznych czynników.
Pod koniec mezozoiku nie było czap polarnych na biegunach, a poziom morza szacunkowo przewyższał dzisiejszy o 100–250 m. Temperatura planety była też znacznie mniej zróżnicowana, bieguny ustępowały terenom równikowym o jedynie 25 °C. Średnia temperatura atmosferyczna była też wyższa od dzisiejszej. Ta ostatnia różnica w przypadku biegunów wynosiła aż 50 °C[165][166].
Skład atmosfery mezozoiku podlega dyskusji. Większość naukowców uważa, że poziom tlenu był znacznie wyższy niż dzisiaj. Inni argumentują, że adaptacje biologiczne odnajdywane u ptaków i innych dinozaurów wskazują na ewolucję w innych warunkach niż w atmosferze bogatej w tlen[167].
Teoria impaktu
edytuj
Teoria impaktu przyciągnęła szerszą, uwagę po pracy z 1980 autorstwa Waltera Alvareza i współpracowników. Wiąże ona wymieranie kredowe z uderzeniem bolidu około 65 milionów lat temu. Naukowcy stwierdzili, że nagły wzrost poziomu irydu odnotowywany na całym świecie w warstwie z tego czasu, stanowić może bezpośredni dowód zderzenia[168]. Siła dowodów wskazuje obecnie, że ciało mierzące od 5 do 10 km średnicy uderzyło w sąsiedztwie Półwyspu Jukatan (obecnie jest to południowy wschód Meksyku). W efekcie utworzył się liczący sobie około 180 km krater Chicxulub oraz nastąpiło masowe wymieranie[169][170]. Naukowcy nie są pewni, czy nieptasie dinozaury zwiększały czy zmniejszały swą liczebność przed zderzeniem. Niektórzy z nich proponują tezę, że meteoryt spowodował nienaturalny wręcz i długo, trwający spadek temperatury atmosfery ziemskiej. Inni uważają, że wywołał raczej niezwykłą falę gorąca. Konsensus wśród wspierających tę teorię badaczy podaje, że uderzenie wywołało wymierania zarówno w sposób bezpośredni (poprzez ciepło na skutek zderzenia), jak i pośredni (poprzez obejmujące całą kulę ziemską ochłodzenie spowodowane materiałem wybitym z krateru, który odbijał promieniowanie słoneczne). Choć nie można określić tempa wymierania na podstawie jedynie zapisu kopalnego, różne modele sugerują, że wymieranie następowało, z niezwykłą prędkością, nawet w ciągu godzin, a nie lat[171].
We wrześniu 2007 amerykańscy badacze pod kierunkiem Williama Bottke z Southwest Research Institute z Boulder (Kolorado) i naukowcy czescy wykorzystali symulacje komputerowe dla identyfikacji prawdopodobnego źródła impaktu Chicxulub. Wyliczyli z prawdopodobieństwem 90%, że asteroida o nazwie Baptistina i średnicy szacunkowo 160 km orbitująca w pasie asteroid rozciągającym się pomiędzy orbitami Marsa i Jowisza została uderzona przez mniejszą, nienazwaną asteroidę o średnicy około 55 km około 160 milionów lat temu. Uderzenie roztrzaskało Baptistinę, tworząc rój istniejący do dziś jako rodzina Baptistina. Obliczenia wskazują, że niektóre fragmenty dostały się na orbity przecinające orbitę Ziemi. Jeden z nich, meteor średnicy 10 km, uderzył w Półwysep Jukatan, tworząc krater Chicxulub[172]. W 2011 nowe dane z Wide-field Infrared Survey Explorer zrewidowały datę kolizji, która zaowocowała powstaniem rodziny Baptistiny na około 80 milionów lat. Czyni to asteroidę tej rodziny wysoce mało prawdopodobną jako przyczynę powstania krateru Chicxulub, jako że procesy rezonansu orbitalnego i kolizji trwają dziesiątki milionów lat[173].
Podobne, ale bardziej kontrowersyjne wyjaśnienie proponuje przejście hipotetycznej gwiazdy Nemezis tworzącej jakoby ze Słońcem układ podwójny przez obłok Oorta, co miało wywołać wiatr komet. Jedna lub więcej z nich miała później uderzyć w Ziemię w mniej więcej tym samym czasie, powodując globalne wymieranie. Jak w przypadku pojedynczej asteroidy, impakt skończył się szybkim spadkiem globalnej temperatury, po którym nastąpił długotrwały okres chłodu[174].
Trapy Dekanu
edytujPrzed 2000 rokiem argumenty mówiące, że pokrywy lawowe trapów Dekanu mają związek z wymieraniem, łączono z poglądem widzącym wymieranie jako proces stopniowy. Uważano bowiem, że tworzenie się trapów rozpoczęło się około 68 milionów lat temu i trwało przez ponad 2 miliony lat. Istnieją jednak dowody, że dwie trzecie trapów ukształtowało się w ciągu miliona lat około 65,5 miliona lat temu. Wobec tego erupcje mogły wywołać naprawdę szybkie wymieranie, możliwe że w czasie tysięcy lat. Cały czas jest to jednak dłużej, niż oczekiwało by się w przypadku pojedynczego zderzenia[175][176].
Trapy Dekanu mogą wiązać się przyczynowo z wymieraniem poprzez kilka mechanizmów. Zalicza się doń wydzielanie do atmosfery pyłu i aerozoli siarkowych, co mogło zablokować dostęp promieni słonecznych i zmniejszyć w ten sposób fotosyntezę roślin. W dodatku wulkanizm związany z trepami Dekanu skutkował emisją do atmosfery dwutlenku węgla, co zwiększyłoby efekt cieplarniany, kiedy atmosfera oczyściła się już z pyłów i aerozoli[176]. Przed masowym wymieraniem dinozaurów wydzielanie gazów wulkanicznych podczas formacji trapów Dekanu przyczyniło się do znacznego globalnego ocieplenia. Niektóre dane skłaniają się ku wzrostowi średniej temperatury o 8 °C w ostatniej połowie miliona lat przed uderzeniem, które spowodowało wytworzenie się krateru Chicxulub[175][176].
W czasie, gdy wiązano trapy Dekanu z wolniejszym wymieraniem, Luis Walter Alvarez, zmarły w 1988, stwierdził, że paleontologów zmyliła mała ilość danych. Jego teza początkowo nie spotkała się z przychylnym przyjęciem. Późniejsze intensywne badania terenowe warstw zawierających skamieliny przydały jego zdaniu większej wagi. W końcu większość paleontologów zaczęło akceptować pogląd, że masowe wymieranie pod koniec kredy wywołane zostało głównie bądź przynajmniej po części przez zderzenie z ciałem niebieskim. Jednakże sam Walter Alvarez przyznał także, że przed impaktem zachodziły na Ziemi inne duże zmiany, jak obniżenie się poziomu morza czy masywne erupcje wulkaniczne, które spowodowały powstaniem trapów Dekanu w Indiach, i zmiany te mogły również przyczynić się do wymierania[177].
Ocaleni
edytujSzczątki nieptasich dinozaurów są okazjonalnie znajdywane ponad granicą kreda-trzeciorzęd. W 2001 paleontolodzy Zielinski i Budahn donieśli o odkryciu pojedynczej kości kończyny hadrozauryda w San Juan Basin w Nowym Meksyku. Opisano ją jako dowód na istnienie paleoceńskich dinozaurów. Formację, w której znaleziono kość, datowano na wczesny paleocen, około 64,5 miliona lat temu, a więc pół miliona lat po rozpoczęciu się kenozoiku[178]. Inne dowody obejmują znalezienie szczątków dinozaurów w formacji Hell Creek aż do 1,3 m powyżej (40 000 lat po) granicy kreda-trzeciorzęd. Podobne doniesienia pochodzą z innych części świata, włączając w to Chiny[179]. Jednak wielu naukowców nie uznaje proponowanych paleoceńskich dinozaurów. Uznają, że ich szczątki opuściły swe pierwotne położenie, po czym zostały pokryte znacznie późniejszymi osadami[180][181]. Jednak bezpośrednie datowanie kości wspiera późniejszy czas ich powstania. Metoda uran-ołów prowadzi do precyzyjnego wyniku 64,8 ± 0,9 miliona lat temu[182]. Jednak nawet jeśli to prawda, garstka nieptasich dinozaurów we wczesnym paleocenie nie zmieni zasadniczych faktów na temat wymarcia[180]
Historia badań
edytujNajwcześniejsze wzmianki
edytujSkamieniałości dinozaurów znano przez tysiąclecia, ale ich prawdziwa natura pozostawała tajemnicą. Chińczycy, którzy dziś określają dinozaury mianem konglong (恐龍, czyli „straszliwy smok”), uważali je za kości smoków i tak też je opisywali. Dla przykładu Hua Yang Guo Zhi, książka napisana przez Zhang Qu podczas panowania dynastii Jin (265-420), donosi o odkryciu kości smoków w Wucheng w prowincji Syczuan[183]. Wieśniacy w centralnych Chinach długo wykorzystywali odsłonięte skamieniałe „kości smoków” w medycynie tradycyjnej, do dziś kontynuują te praktyki[184]. W Europie ogólnie uważano skamieniałości dinozaurów za szczątki gigantów i innych stworzeń zabitych przez potop.

Opisy naukowe znalezisk uznanych później za kości dinozaurów pojawiają się po raz pierwszy w siedemnastowiecznej Anglii. Fragment kości, obecnie zidentyfikowany jako część kości udowej megalozaura[185], wydobyto z kopalni wapienia w Cornwell w okolicy Chipping Norton w Oxfordshire w Anglii w 1676. Wysłano go Robertowi Plotowi, profesorowi chemii na Uniwersytecie Oksfordzkim i pierwszemu kustoszowi Ashmolean Museum, który opublikował opis znaleziska w Natural History of Oxfordshire w 1677. Poprawnie określił on je jako dystalny koniec kości udowej dużego zwierzęcia, stwierdził też, że jest za duży, by należeć do jakiegokolwiek znanego wtedy gatunku. Doszedł jednak do wniosku, że chodzi o kość udową wielkiego człowieka podobnego do wymienionych w Biblii. W 1699 Edward Lhuyd, przyjaciel Isaaca Newtona, po raz pierwszy potraktował naukowo znalezisko określane obecnie jako ząb zauropoda, „Rutellum implicatum”[186][187], który znaleziono w Caswell w pobliżu Witney, Oxfordshire[188].
Pierwsze badania naukowe
edytuj
Pomiędzy 1815 i 1824 William Buckland, profesor geologii na Uniwersytecie Oksfordzkim, zebrał więcej skamieniałych szczątków megalozaura, został też pierwszą osobą, która opisała dinozaura w czasopiśmie naukowym[185][189]. Drugim takim rodzajem był Iguanodon, odnaleziony w 1822 przez Mary Ann Mantell – żonę angielskiego geologa Gideona Mantella. Gideon Mantell dopatrzył się podobieństw pomiędzy tymi szczątkami a należącymi do współczesnego legwana. Opublikował swe odkrycie w 1825[190][191].
Badania „wielkich, skamieniałych jaszczurów” szybko stały się obiektem wielkiego zainteresowania naukowców europejskich i amerykańskich. W 1842 angielski paleontolog Richard Owen stworzył termin „dinozaury”. Stwierdził, że znalezione dotąd szczątki iguanodonta, megalozaura i hyleozaura dzielą liczne wyróżniające się cechy, dlatego też zdecydował się połączyć je w odrębny takson. Za wsparciem księcia Alberta, męża królowej Wiktorii, Owen założył Muzeum Historii Naturalnej w Londynie w South Kensington w Londynie, by wystawiać narodową kolekcję dinozaurzych skamielin, jak też prowadzić inne wystawy geologiczne i biologiczne[192].


„Manię” dinozaurów obrazuje zażarta rywalizacja pomiędzy Edwardem Drinkerem Cope’em i Othnielem Charlesem Marshem. Obaj paleontolodzy starali się być pierwszymi, którzy znajdą nowe dinozaury. Ich zmagania stały się znane jako wojny o kości. Konflikt prawdopodobnie rozpoczął się, gdy Marsh publicznie wytknął Cope’owi błąd w rekonstrukcji elasmozaura (Cope pomyłkowo umieścił głowę plezjozaura na końcu jego ogona). Walka między badaczami trwała 30 lat. Zakończyła się w 1897 śmiercią Cope’a, po przeznaczeniu całej swej fortuny na poszukiwania dinozaurów. Marsh wygrał, głównie dzięki powiązaniom z US Geological Survey. Niestety wiele wartościowych okazów dinozaurów zostało uszkodzonych lub zniszczonych przez drastyczne metody stosowane przez poszukiwaczy. Na przykład używano dynamitu do wydobywania kości, co dzisiaj paleontolodzy uważają za bulwersujące. Pomimo prymitywnych metod Cope i Marsh bardzo przyczynili się do rozwoju paleontologii. Marsh odkrył 86 nowych gatunków dinozaurów, podczas gdy Cope – 56, w sumie 142 nowe gatunki. Kolekcja Cope’a znajduje się obecnie w Amerykańskim Muzeum Historii Naturalnej w Nowym Jorku, natomiast okazy Marsha wystawia Peabody Museum of Natural History, należące do Uniwersytetu Yale[193].
Odrodzenie dinozaurów
edytuj
Na polu badań dinozaurów aktywność zwiększała się znacznie od lat 60. XX wieku i dzieje się tak do tej pory. Po części zapoczątkował to John Ostrom swoim odkryciem deinonycha, aktywnego drapieżnika, który mógł być ciepłokrwisty. Kontrastowało to z rozpowszechnionym wcześniej wizerunkiem dinozaurów jako stworzeń powolnych, zimnokrwistych. Paleontologia kręgowców stała się nauką globalną. Nowe odkrycia dinozaurów poczyniono na obszarach wcześniej nieeksploatowanych pod tym względem, jak Indie, Ameryka Południowa, Madagaskar, Antarktyka, a w największym stopniu Chiny (niezwykle dobrze zachowane upierzone dinozaury z Chin wsparły później istnienie powiązania pomiędzy nieptasimi dinozaurami i ich dzisiejszymi krewnymi, ptakami). Szerokie zastosowanie kladystyki, rygorystycznie analizującej pokrewieństwo pomiędzy organizmami, stało się bardzo użyteczne w klasyfikowaniu dinozaurów. Analiza kladystyczna razem z innymi nowoczesnymi technikami pomogła poradzić sobie z często niekompletnym, fragmentarycznym zapisem kopalnym[194].
Tkanki miękkie i DNA
edytujJeden z najlepszych przykładów odcisków tkanek miękkich na skamielinach dinozaurów stanowi odkrycie z Pietraroja we Włoszech. Odkrycie odnotowano w 1998. Chodziło o okaz małego, bardzo młodego celurozaura Scipionyx samniticus. Znalezisko obejmuje fragmenty jelit, w tym okrężnicy, wątroby, mięśni i tchawicy młodego dinozaura[82].
W marcu 2005 w publikacji w Science paleontolog Mary Higby Schweitzer i jej zespół ogłosili odkrycie giętkiego materiału przypominającego tkankę miękką w obrębie liczącej 68 milionów lat kości kończyny dolnej Tyrannosaurus rex z montańskiej formacji Hell Creek. Po odkryciu tkanka została uwodniona przez zespół badawczy[83].
Po kilkutygodniowych pracach w celu usunięcia minerałów ze skamieniałej jamy szpikowej w procesie demineralizacji Schweitzer znalazła oznaki istnienia nietkniętych struktur, jak naczynia krwionośne, osteony i tkanka łączna tworząca włókna. Analiza mikroskopowa ujawniła następnie, że domniemana tkanka miękka dinozaura zachowała drobne struktury, nawet na poziomie komórkowym. Właściwa natura i kompozycja tego materiału, jak też implikacje odkrycia zespołu Schweitzer nie są jeszcze całkiem jasne[83].
W 2009 zespół obejmujący Schweitzer ogłosił powtórzenie tych wyników przy użyciu bardziej skrupulatnej metodologii. Tym razem znaleziono podobną tkankę miękką kaczodziobego Brachylophosaurus canadensis z montańskiej formacji Judith River. Materiał obejmował więcej szczegółów tkanki, w tym komórki zdające się nawet posiadać pozostałości jąder oraz coś, co wydawało się erytrocytami. Inny materiał z tych kości zawierał kolagen, jak i we wspomnianej powyżej kości tyranozaura. Typ wytwarzanego przez zwierzę kolagenu odpowiadał spotykanemu u współczesnego kurczaka czy strusia[195].
W dwóch przypadkach doniesiono nawet o skutecznej ekstrakcji DNA z dinozaurzej skamieliny. Jednak dalsze sprawdzanie i procedura peer review nie potwierdziły żadnego z nich[196]. Jednak udało się wywnioskować budowę peptydu zaangażowanego w widzenie teoretycznego dinozaura dzięki metodom rekonstrukcji filogenetycznych na bazie sekwencjonowania DNA blisko spokrewnionych zwierząt współczesnych, jak gady czy ptaki[197]. W dodatku pewne białka, w tym hemoglobina[198], zostały wstępnie wykryte w skamielinach dinozaurów[199][200].
Czas odkrycia ważniejszych rodzajów
edytuj|
Czas odkrycia ważniejszych rodzajów |
|---|
 |
W kulturze
edytuj
Dla człowieka dinozaury (nieptasie) były stworzeniami o fantastycznym wyglądzie i często olbrzymich rozmiarach. W takiej formie istnieją w popularnych wyobrażeniach, stając się trwałą częścią kultury. Pojawienie się słowa „dinozaur” w języku potocznym odzwierciedla ich znaczenie, np. w języku angielskim słowem tym określa się coś niepraktycznie wielkiego, przestarzałego, zmierzającego ku zagładzie[201].

Entuzjazm opinii publicznej wobec tych imponujących zauropsydów wystąpił po raz pierwszy w Anglii epoki wiktoriańskiej. Właśnie tam w 1854, trzy dekady po pierwszych naukowych opisach szczątków dinozaurów, wystawiono w Crystal Palace Park w Londynie rzeźby przedstawiające te stworzenia. Dinozaury z Crystal Palace stały się tak popularne, że w niedługim czasie rozwinął się silny rynek kopii. W kolejnych dekadach wystawy przedstawiające te zwierzęta otwierano w parkach i muzeach wielu krajów[202]. Trwała popularność dinozaurów zaowocowała znacznym publicznym wsparciem nauki badającej te kręgowce, a w konsekwencji nowymi odkryciami. Dla przykładu współzawodnictwo pomiędzy muzeami w Stanach Zjednoczonych o przyciągnięcie opinii publicznej doprowadziło do wojny o kości w dwóch ostatnich dekadach XIX wieku, podczas której dwóch zwaśnionych paleontologów starało się o zdobycie pierwszeństwa w liczbie opisanych rodzajów[203].
Wcześniejsza obecność dinozaurów w kulturze masowej zapewniła im miejsce w literaturze, filmie i innych mediach. Począwszy od 1852 (wzmianka w Bleak House Karola Dickensa)[204] przedstawiano te stworzenia w wielkiej liczbie dzieł. Juliusz Verne wydał w 1864 powieść Podróż do wnętrza Ziemi, Arthur Conan Doyle w 1912 – książkę Zaginiony świat. Inne przykłady to film z 1933 King Kong, powstały w 1954 Godzilla i wiele jego sequeli, bestseller z 1990 Park Jurajski autorstwa Michaela Crichtona oraz jego adaptacja filmowa z 1993. Autorzy prac na temat dinozaurów, w tym pewni prominentni paleontolodzy, często wykorzystywali dinozaury, by uczyć o nauce w ogólności. Dinozaury są też powszechnie spotykane w reklamie. Liczne firmy posłużyły się nimi w reklamówkach drukowanych bądź telewizyjnych, by sprzedać własne produkty bądź przedstawić konkurentów jako wolnych, mgliście myślących czy przestarzałych[205].
Uwagi
edytujPrzypisy
edytuj- ↑ a b c d Alan Feduccia, Birds are dinosaurs: simple answer to a complex problem, „The Auk”, 119 (4), 2002, s. 1187–1201, DOI: 10.1642/0004-8038(2002)119[1187:BADSAT]2.0.CO;2.
- ↑ a b c (1) Thomas R. Holtz Jr., Luis V. Rey: Dinosaurs: the most complete, up-to-date encyclopedia for dinosaur lovers of all ages. New York: Random House, 2007. ISBN 0-375-82419-7. (2) Thomas R. Holtz Jr. (2011) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2011 Appendix.
- ↑ John R. Hutchinson: Are Birds Really Dinosaurs?. University of California Museum of Paleontology, 1998-01-22. [dostęp 2017-04-09]. (ang.).
- ↑ Nicholas St. Fleur: That Thing With Feathers Trapped in Amber? It Was a Dinosaur Tail. 8 grudnia 2016. (ang.).
- ↑ Michael S.Y. Lee, Andrea Cau, Darren Naish, Gareth J. Dyke. Sustained miniaturization and anatomical innovation in the dinosaurian ancestors of birds. „Science”. 345 (6196), s. 562–566, 1 sierpnia 2014. DOI: 10.1126/science.1252243. PMID: 25082702. Bibcode: 2014Sci...345..562L. [dostęp 2014-08-02]. (ang.).
- ↑ Thomas R. Holtz Jr.: Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. Random House, 2007. ISBN 0-375-82419-7. (ang.).
- ↑ Bradley C. Livezey, Richard L. Zusi. Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy. II. Analysis and discussion. „Zoological Journal of the Linnean Society”. 149, s. 1–95, 2007. DOI: 10.1111/j.1096-3642.2006.00293.x. (ang.).
- ↑ Luis M. Chiappe, Ji Shu’an, Ji Qiang: Anatomy and systematics of the Confuciusornithidae (Theropoda: Aves) from the Late Mesozoic of northeastern China. W: Bulletin-American Museum of Natural History. American Museum of Natural History, 1999. ISSN 0003-0090.
- ↑ Luis M. Chiappe, John R. Hutchinson. The First Known Alvarezsaurid (Theropoda: Aves) from North America. „Journal of Vertebrate Paleontology”. 18, s. 447–450, 1998. DOI: 10.1080/02724634.1998.10011073. (ang.).
- ↑ Mark A. Norell, Qiang Ji, Keqin Gao, Chongxi Yuan, Yibin Zhao, Lixia Wang. Palaeontology: ‘Modern’ feathers on a non-avian dinosaur. „Nature”. 416, s. 36–37, 2002. DOI: 10.1038/416036a. (ang.).
- ↑ Xing Xu, Zhonghe Zhou, Xiaolin Wang. The smallest known non-avian theropod dinosaur. „Nature”. 408, s. 705–708, 2000. DOI: 10.1038/35047056. (ang.).
- ↑ Xu Xing, Mark A. Norell. Non-avian dinosaur fossils from the Lower Cretaceous Jehol Group of western Liaoning, China. „Geological Journal”. 41, s. 419–437, 2006. DOI: 10.1002/gj.1044. (ang.).
- ↑ David C. Evans. Nowe dane o zależności między mózgiem a puszką mózgową u dinozaurów ptasiomiednicznych. „Acta Palaeontologica Polonica”. 50, s. 617–622, 2005.
- ↑ Magdalena Borsuk-Białynicka. Dinozaury w nauce polskiej. „Ewolucja. Biuletyn Muzeum Ewolucji Instytutu Paleobiologii PAN”. 3, s. 28–34, 2008. Muzeum Ewolucji Instytutu Paleobiologii PAN.
- ↑ Numbers of threatened species by major groups of organisms (1996–2012) [online], IUCN Red List, 2012 [zarchiwizowane z adresu 2013-02-04].
- ↑
 Michael E. Alfaro, Francesco Santini, Chad Brock, Hugo Alamillo i inni. Nine exceptional radiations plus high turnover explain species diversity in jawed vertebrates. „Proceedings of the National Academy of Sciences”. 106 (32), s. 13410–13414, 2009. DOI: 10.1073/pnas.0811087106. PMID: 19633192. PMCID: PMC2715324. Bibcode: 2009PNAS..10613410A.
Michael E. Alfaro, Francesco Santini, Chad Brock, Hugo Alamillo i inni. Nine exceptional radiations plus high turnover explain species diversity in jawed vertebrates. „Proceedings of the National Academy of Sciences”. 106 (32), s. 13410–13414, 2009. DOI: 10.1073/pnas.0811087106. PMID: 19633192. PMCID: PMC2715324. Bibcode: 2009PNAS..10613410A.
- ↑ a b c Steve C. Wang, Peter Dodson. Estimating the Diversity of Dinosaurs. „Proceedings of the National Academy of Sciences”. 103 (37), s. 13601–13605, 2006. DOI: 10.1073/pnas.0606028103. PMID: 16954187. PMCID: PMC1564218. Bibcode: 2006PNAS..10313601W.
- ↑ a b Jonathan Amos: Will the real dinosaurs stand up?. [w:] BBC News [on-line]. 2008-09-17. [dostęp 2011-03-23].
- ↑ Dinosaur Genera List. [dostęp 2014-04-12]. (ang.).
- ↑ a b c N. MacLeod, P.F. Rawson, P.L. Forey, F.T. Banner i inni. The Cretaceous–Tertiary biotic transition. „Journal of the Geological Society”. 154 (2), s. 265–292, 1997. DOI: 10.1144/gsjgs.154.2.0265.
- ↑ a b c d e f g h Kenneth Carpenter: Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus. W: John R. Foster, Spencer G. Lucas: Paleontology and Geology of the Upper Jurassic Morrison Formation. Albuquerque: New Mexico Museum of Natural History and Science, 2006, s. 131–138, seria: New Mexico Museum of Natural History and Science Bulletin 36.
- ↑ a b Owen 1842 ↓, s. 103.
- ↑ Liddell–Scott–Jones Lexicon of Classical Greek. [dostęp 2008-08-05].
- ↑ James O. Farlow, M.K. Brett-Surman: Preface. W: James O. Farlow, M.K. Brett-Surman: The Complete Dinosaur. Indiana University Press, 1997, s. ix–xi. ISBN 0-253-33349-0.
- ↑ a b c d e Michael J. Benton: Origin and Relationships of Dinosauria. W: David B. Weishampel, Peter Dodson, Halszka Osmólska: The Dinosauria (2nd edition). Berkeley: University of California Press, 2004, s. 7–19. ISBN 0-520-24209-2.
- ↑ George Olshevsky. An annotated checklist of dinosaur species by continent. „Mesozoic Meanderings”. 3, s. 1–157, 2000.
- ↑ Paul C. Sereno. The logical basis of phylogenetic taxonomy. „Systematic Biology”. 54 (4), s. 595–619, 2005. DOI: 10.1080/106351591007453. PMID: 16109704.
- ↑ a b Matthew G. Baron, David B. Norman, Paul M. Barrett. A new hypothesis of dinosaur relationships and early dinosaur evolution. „Nature”. 543 (7646), s. 501–506, 2017. DOI: 10.1038/nature21700. (ang.).
- ↑ Dinosauria R. Owen 1842 [M.C. Langer, F.E. Novas, J.S. Bittencourt, M. D. Ezcurra, and J.A. Gauthier], converted clade name. W: Phylonyms. A Companion to the PhyloCode. Kevin de Queiroz, Philip D. Cantino i Jacques A. Gauthier (red.). Boca Raton: CRC Press, 2020. ISBN 978-0-429-44627-6.
- ↑ Donald F. Glut: Dinosaurs: The Encyclopedia. Jefferson, North Carolina: McFarland & Co, 1997, s. 40. ISBN 0-89950-917-7.
- ↑ David Lambert: The Dinosaur Data Book. and the Diagram Group. New York: Avon Books, 1990, s. 288. ISBN 0-380-75896-2.
- ↑ Michael Morales: Nondinosaurian Vertebrates of the Mesozoic. W: James O. Farlow, M.K. Brett-Surman: The Complete Dinosaur. Bloomington: Indiana University Press, 1997, s. 607–624. ISBN 0-253-33349-0.
- ↑ George Olshevsky: Dinosaur Genera List. [dostęp 2011-11-24]. (ang.).
- ↑ Dale A. Russell. China and the lost worlds of the dinosaurian era. „Historical Biology”. 10, s. 3–12, 1995. DOI: 10.1080/10292389509380510.
- ↑ Romain Amiot, Eric Buffetaut, Christophe Lécuyer, Xu Wang i inni. Oxygen isotope evidence for semi-aquatic habits among spinosaurid theropods. „Geology”. 38 (2), s. 139–142, 2010. DOI: 10.1130/G30402.1.
- ↑ Sterling J. Nesbitt. The early evolution of archosaurs. Relationships and the origin of major clades. „Bulletin of the American Museum of Natural History”. 352, s. 1–292, 2011. DOI: 10.1206/352.1.
- ↑ a b Thomas R. Holtz Jr: Classification and evolution of the dinosaur groups. W: Gregory S. Paul: The Scientific American Book of Dinosaurs. St. Martin’s Press, 2000, s. 140–168. ISBN 0-312-26226-4.
- ↑ Dave Smith, Dinosauria Morphology, UC Berkeley [dostęp 2013-01-21].
- ↑ Max C. Langer, Fernando Abdala, Martha Richter, Michael J. Benton. A sauropodomorph dinosaur from the Upper Triassic (Carnian) of southern Brazil. „Comptes Rendus de l’Academie des Sciences, Paris: Sciences de la terre et des planètes”. 329 (7), s. 511–517, 1999. DOI: 10.1016/S1251-8050(00)80025-7. Bibcode: 1999CRASE.329..511L.
- ↑ Martín D. Ezcurra. A new early dinosaur (Saurischia: Sauropodomorpha) from the Late Triassic of Argentina: a reassessment of dinosaur origin and phylogeny. „Journal of Systematic Palaeontology”. 8 (3), s. 371–425, 2010. DOI: 10.1080/14772019.2010.484650. (ang.).
- ↑ Sterling J. Nesbitt, Randall B. Irmis, William G. Parker. A critical re-evaluation of the Late Triassic dinosaur taxa of North America. „Journal of Systematic Palaeontology”. 5 (2), s. 209–243, 2007. DOI: 10.1017/S1477201907002040.
- ↑ a b c d e Tom Hogervorst, Heinse W. Bouma, John de Vos. Evolution of the hip and pelvis. „Acta Orthopaedica”. 80, s. 1–39, 2009. DOI: 10.1080/17453690610046620. (ang.).
- ↑ Co odkryto nie później, niż w 1909: Dr. Holland and the Sprawling Sauropods [online], hmnh.org [zarchiwizowane z adresu 2011-06-12]. Argumentację i rysunki prezentuje również Adrian Desmond: Hot Blooded Dinosaurs. DoubleDay, 1976. ISBN 0-385-27063-1.
- ↑ Adrian Desmond: Hot Blooded Dinosaurs. DoubleDay, 1976. ISBN 0-385-27063-1.
- ↑ Dinosaurs. W: Richard Cowen: History of Life. Wyd. 4. Blackwell Publishing, 2004, s. 151–175. ISBN 1-4051-1756-7. OCLC 53970577.
- ↑ a b Tai Kubo, Michael J. Benton. Evolution of hindlimb posture in archosaurs: limb stresses in extinct vertebrates. „Palaeontology”. 50 (6), s. 1519–1529, 2007. DOI: 10.1111/j.1475-4983.2007.00723.x.
- ↑ Lee R. Kump, Alexander Pavlov, Michael A. Arthur. Massive release of hydrogen sulfide to the surface ocean and atmosphere during intervals of oceanic anoxia. „Geology”. 33 (5), s. 397–400, 2005. DOI: 10.1130/G21295.1. Bibcode: 2005Geo....33..397K.
- ↑ L.H. Tanner, S.G. Lucas, M.G. Chapman. Assessing the record and causes of Late Triassic extinctions. „Earth-Science Reviews”. 65 (1–2), s. 103–139, 2004. DOI: 10.1016/S0012-8252(03)00082-5. Bibcode: 2004ESRv...65..103T.
- ↑ a b Paul C. Sereno. The evolution of dinosaurs. „Science”. 284 (5423), s. 2137–2147, 1999. DOI: 10.1126/science.284.5423.2137. PMID: 10381873.
- ↑ Paul C. Sereno, Catherine A. Forster, Raymond R. Rogers, Alfredo M. Monetta. Primitive dinosaur skeleton from Argentina and the early evolution of Dinosauria. „Nature”. 361 (6407), s. 64–66, 1993. DOI: 10.1038/361064a0. Bibcode: 1993Natur.361...64S.
- ↑ Ricardo N. Martinez. A Basal Dinosaur from the Dawn of the Dinosaur Era in Southwestern Pangaea. „Science”. 331 (6014), s. 206–210, 2011. (ang.).
- ↑ Sterling J. Nesbitt, Paul M. Barrett, Sarah Werning, Christian A. Sidor, Alan J. Charig. The oldest dinosaur? A Middle Triassic dinosauriform from Tanzania. „Biology Letters”. 9 (1), s. 20120949, 2013. Royal Society Publishing. DOI: 10.1098/rsbl.2012.0949. (ang.).
- ↑ Sterling J. Nesbitt. The early evolution of archosaurs: relationships and the origin of major clades. „Bulletin of the American Museum of Natural History”, s. 1–292, 2011. DOI: 10.1206/352.1. (ang.).
- ↑ Stephen L. Brusatte, Michael J. Benton, Marcello Ruta, Graeme T. Lloyd. Superiority, Competition, and Opportunism in the Evolutionary Radiation of Dinosaurs. „Science”. 321, s. 1485–1488, 2008-09-12. (ang.).
- ↑ a b c d Thomas R Holtz Jr., Ralph E. Chapman, Matthew C. Lamanna: Mesozoic Biogeography of Dinosauria. W: David B. Weishampel, Peter Dodson, Halszka Osmólska: The Dinosauria (2nd edition). Berkeley: University of California Press, 2004, s. 627–642. ISBN 0-520-24209-2.
- ↑ a b c d e David E. Fastovsky, Joshua B. Smith: Dinosaur Paleoecology. W: David B. Weishampel, Peter Dodson, Halszka Osmólska: The Dinosauria (2nd edition). Berkeley: University of California Press, 2004, s. 614–626. ISBN 0-520-24209-2.
- ↑ Octávio Mateus. Lourinhanosaurus antunesi, A New Upper Jurassic Allosauroid (Dinosauria: Theropoda) from Lourinhã, Portugal. „Memórias da Academia de Ciências de Lisboa”. 37, s. 111–124, 1998. (ang.).
- ↑ a b c d e f Kevin Padian: Basal Avialae. W: The Dinosauria (2d edition). David B. Weishampel, Peter Dodson, Halszka Osmólska. University of California Press, 2004, s. 210–231. ISBN 0-520-24209-2.
- ↑ a b David J. Varricchio, Anthony J. Martin, Yoshihiro Katsura. First trace and body fossil evidence of a burrowing, denning dinosaur. „Proceedings of the Royal Society B: Biological Sciences”. 274 (1616), s. 1361–1368, 2007. DOI: 10.1098/rspb.2006.0443. PMID: 17374596. PMCID: PMC2176205.
- ↑ Paul C. Sereno, Jeffrey A. Wilson, Lawrence M. Witmer, John A. Whitlock i inni. Structural extremes in a Cretaceous dinosaur. „PLoS ONE”. 2 (11), s. e1230, 2007. DOI: 10.1371/journal.pone.0001230. PMID: 18030355. PMCID: PMC2077925. Bibcode: 2007PLoSO...2.1230S.
- ↑ Vandana Prasad, Caroline A.E. Strömberg, Habib Alimohammadian, Ashok Sahni. Dinosaur coprolites and the early evolution of grasses and grazers. „Science”. 310 (5751), s. 1170–1180, 2005. DOI: 10.1126/science.1118806. PMID: 16293759. Bibcode: 2005Sci...310.1177P.
- ↑ J. David Archibald, David E. Fastovsky: Dinosaur Extinction. W: David B. Weishampel, Peter Dodson, Halszka Osmólska: The Dinosauria (2d edition). Berkeley: University of California Press, 2004, s. 672–684. ISBN 0-520-24209-2.
- ↑ Bent E.K. Lindow, Bird Evolution Across the K–Pg Boundary and the Basal Neornithine Diversification, [w:] Gareth Dyke, Gary Kaiser, Living Dinosaurs: The Evolutionary History of Modern Birds, Chichester, UK: John Wiley & Sons, Ltd, 2011, s. 338–354, DOI: 10.1002/9781119990475.ch14, ISBN 978-1-119-99047-5 (ang.).
- ↑ Michael J. Benton: Vertebrate Paleontology. Blackwell Publishers, 2004, s. xii–452. ISBN 0-632-05614-2.
- ↑ Gregory S. Paul: Predatory Dinosaurs of the World. New York: Simon and Schuster, 1988, s. 248–250. ISBN 0-671-61946-2.
- ↑ James M. Clark, Teresa Maryańska, Rinchen Barsbold: Therizinosauroidea. W: David B. Weishampel, Peter Dodson, Halszka Osmólska: The Dinosauria (2nd edition). University of California Press, 2004, s. 151–164. ISBN 0-520-24209-2.
- ↑ Mark A. Norell, Peter J. Makovicky: Dromaeosauridae. W: David B. Weishampel, Peter Dodson, Halszka Osmólska: The Dinosauria (2d edition). University of California Press, 2004, s. 196–210. ISBN 0-520-24209-2.
- ↑ David E. Fastovsky, David B. Weishampel: Dinosaurs A Concise Natural History. 2013, s. 70. ISBN 978-1-107-27309-2.
- ↑ David E. Fastovsky, David B. Weishampel: Dinosaurs A Concise Natural History. 2013, s. 75–76. ISBN 978-1-107-27309-2.
- ↑ Andrea Cau. The assembly of the avian body plan: a 160-million-year long process. „Bollettino della Società Paleontologica Italiana”. 57 (1), s. 1–25, 2018. DOI: 10.4435/BSPI.2018.01. (ang.).
- ↑ Max C. Langer, Martín D. Ezcurra, Oliver W.M. Rauhut, Michael J. Benton, Fabien Knoll, Blair W. McPhee, Fernando E. Novas, Diego Pol i Stephen L. Brusatte. Untangling the dinosaur family tree. „Nature”. 551 (7678), s. E1–E3, 2017. DOI: 10.1038/nature24011. (ang.).
- ↑ a b c Henryk Szarski: Historia zwierząt kręgowych. Wyd. V zmienione. Warszawa: PWN, 1990, s. 567. ISBN 83-01-09155-X.
- ↑ a b Douglas J. Futuyma: Ewolucja. Wyd. 1. Warszawa: Wydawnictwa Uniwersytetu Warszawskiego, 2008, s. 606. ISBN 978-83-235-0577-8.
- ↑ a b Richard O. Prum. Why ornitologists should care about the theropod origins of birds. „The Auk”. 119 (1), s. 1–17, 2002. The American Ornithologists’ Union. DOI: 10.1642/0004-8038(2002)119[0001:WOSCAT]2.0.CO;2.
- ↑ Darren Naish: Birds. W: M.K. Brett-Surman, Thomas R. Holtz, Jr, James O. Farlow: The Complete Dinosaur. Wyd. drugie. Bloomington: Indiana University Press, 2012, s. 379–423. ISBN 978-0-253-35701-4.
- ↑ Classification of the vertebrates, [w:] Michael J. Benton, Vertebrate paleontology, wyd. 3, University of Bristol, 2004, ISBN 978-0-632-05637-8 [zarchiwizowane z adresu 2018-10-15].
- ↑ Aves, [w:] Integrated Taxonomic Information System [dostęp 2014-11-14] (ang.).
- ↑ vertebrate, [w:] Encyclopædia Britannica [dostęp 2017-04-22] (ang.).
- ↑ E. Connolly: Introduction to Zoology. [w:] Moorpark College D.V.M. [on-line]. 2013. [dostęp 2014-11-14]. [zarchiwizowane z tego adresu (2014-11-29)]. (ang.).
- ↑ Peter M. Galton, On Staurikosaurus pricei, an early saurischian dinosaur from the Triassic of Brazil, with notes on the Herrerasauridae and Poposauridae, „Paläontologische Zeitschrift”, 51 (3–4), 1977, s. 234–245, DOI: 10.1007/BF02986571 (ang.).
- ↑ Davide Pisani i inni, A genus-level supertree of the Dinosauria, „Proceedings of the Royal Society B”, 269 (1494), 2002, s. 915–921, DOI: 10.1098/rspb.2001.1942, PMID: 12028774, PMCID: PMC1690971 (ang.).
- ↑ a b Cristiano Dal Sasso, Marco Signore. Exceptional soft-tissue preservation in a theropod dinosaur from Italy. „Nature”. 392 (6674), s. 383–387, 1998. DOI: 10.1038/32884. Bibcode: 1998Natur.392..383D.
- ↑ a b c Mary H. Schweitzer, Jennifer L. Wittmeyer, John R. Horner, Jan K. Toporski. Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus rex. „Science”. 307 (5717), s. 1952–1955, 2005. DOI: 10.1126/science.1108397. PMID: 15790853. Bibcode: 2005Sci...307.1952S.
- ↑ James O. Farlow. On the rareness of big, fierce animals: speculations about the body sizes, population densities, and geographic ranges of predatory mammals and large, carnivorous dinosaurs. „American Journal of Science”. 293-A, s. 167–199, styczeń 1993. DOI: 10.2475/ajs.293.A.167.
- ↑ Jan Peczkis. Implications of body-mass estimates for dinosaurs. „Journal of Vertebrate Paleontology”. 14 (4), s. 520–533, 1994. DOI: 10.1080/02724634.1995.10011575.
- ↑ Anatomy and evolution [online], National Museum of Natural History [zarchiwizowane z adresu 2009-04-07].
- ↑ P. Martin Sander, Andreas Christian, Marcus Clauss, Regina Fechner i inni. Biology of the sauropod dinosaurs: the evolution of gigantism. „Biological Reviews”. 86 (1), s. 117–155, 2011. DOI: 10.1111/j.1469-185X.2010.00137.x. PMID: 21251189. PMCID: PMC3045712.
- ↑ a b c d Gary P. Burness, Timothy Flannery, Jared Diamond. Dinosaurs, dragons, and dwarfs: The evolution of maximal body size. „Proceedings of the National Academy of Sciences”. 98 (25), s. 14518–14523, 2001. DOI: 10.1073/pnas.251548698. PMID: 11724953. Bibcode: 2001PNAS...9814518B. (ang.).
- ↑ a b c Jürgen Hummel, Marcus Clauss: Sauropod feeding and digestive physiology. W: Nicole Klein, Kristian Remes, Carole T. Gee, P. Martin Sander: Biology of the sauropod dinosaurs: Understanding the life of giants. Indiana University Press, 2011, s. 11–33. ISBN 0-253-35508-7.
- ↑ Edwin Harris Colbert: Men and dinosaurs: the search in field and laboratory. Harmondsworth: Penguin, 1971. ISBN 0-14-021288-4. (ang.).
- ↑ a b Gerardo V. Mazzetta, Per Christiansen, Richard A. Fariña. Giants and bizarres: body size of some southern South American Cretaceous dinosaurs. „Historical Biology”. 2004, s. 1–13, 2004. (ang.).
- ↑ David M. Lovelace. Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny. „Arquivos do Museu Nacional”. 65 (4), s. 527–544, 2007.
- ↑ Cristiano dal Sasso i inni, New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities, „Journal of Vertebrate Paleontology”, 25 (4), 2006, s. 888–896, DOI: 10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2.
- ↑ François Therrien, Donald M. Henderson. My theropod is bigger than yours ... or not: estimating body size from skull length in theropods. „Journal of Vertebrate Paleontology”. 27 (1), s. 108–115, 2007. DOI: 10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2.
- ↑ a b Fucheng Zhang, Zhonghe Zhou, Xing Xu, Xiaolin Wang i inni. A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers. „Nature”. 455 (7216), s. 1105–1108, 2008. DOI: 10.1038/nature07447. PMID: 18948955. Bibcode: 2008Natur.455.1105Z.
- ↑ a b Xing Xu, Qi Zhao, Mark A. Norell, Corwin Sullivan i inni. A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin. „Chinese Science Bulletin”. 54 (3), s. 430–435, 2008. DOI: 10.1007/s11434-009-0009-6.
- ↑ R.J. Butler, Q. Zhao. The small-bodied ornithischian dinosaurs Micropachycephalosaurus hongtuyanensis and Wannanosaurus yansiensis from the Late Cretaceous of China. „Cretaceous Research”. 30 (1), s. 63–77, 2009. DOI: 10.1016/j.cretres.2008.03.002.
- ↑ Johan Yans, Jean Dejax, Denise Pons, Christian Dupuis i inni. Implications paléontologiques et géodynamiques de la datation palynologique des sédiments à faciès wealdien de Bernissart (bassin de Mons, Belgique). „Comptes Rendus Palevol”. 4 (1–2), s. 135–150, 2005. DOI: 10.1016/j.crpv.2004.12.003. (fr.).
- ↑ David B. Norman. A mass-accumulation of vertebrates from the Lower Cretaceous of Nehden (Sauerland), West Germany. „Proceedings of the Royal Society of London. Series B, Biological Sciences”. 230 (1259), s. 215–255, marzec 1987. DOI: 10.1098/rspb.1987.0017.
- ↑ Julia J. Day, Paul Upchurch, David B. Norman, Andrew S. Gale i inni. Sauropod trackways, evolution, and behavior. „Science”. 296 (5573), s. 1659, 2002. DOI: 10.1126/science.1070167. PMID: 12040187.
- ↑ Joanna L. Wright: Steps in Understanding Sauropod Biology. W: Kristina A. Curry Rogers, Jeffrey A. Wilson: The Sauropods: Evolution and Paleobiology. Berkeley: University of California Press, 2005, s. 252–284. ISBN 0-520-24623-3.
- ↑ David J. Varricchio, Paul C. Sereno, Zhao Xijin, Tan Lin i inni. Mud-trapped herd captures evidence of distinctive dinosaur sociality. „Acta Palaeontologica Polonica”. 53 (4), s. 567–578, 2008. DOI: 10.4202/app.2008.0402. [dostęp 2011-05-06].
- ↑ Allosaurus. W: Don Lessem, Donald F. Glut: The Dinosaur Society’s Dinosaur Encyclopedia. Random House, 1993, s. 19–20. ISBN 0-679-41770-2.
- ↑ W. Desmond Maxwell, John H. Ostrom. Taphonomy and paleobiological implications of Tenontosaurus–Deinonychus associations. „Journal of Vertebrate Paleontology”. 15 (4), s. 707–712, 1995. DOI: 10.1080/02724634.1995.10011256.
- ↑ Brian T. Roach, Daniel L. Brinkman. A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs. „Bulletin of the Peabody Museum of Natural History”. 48 (1), s. 103–138, 2007. DOI: 10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2.
- ↑ Darren Tanke, Head-biting behavior in theropod dinosaurs: paleopathological evidence, „Gaia”, 15, 1998, s. 167–184 [zarchiwizowane z adresu 2008-02-27].
- ↑ The Fighting Dinosaurs [online], American Museum of Natural History [zarchiwizowane z adresu 2012-01-18].
- ↑ a b Kenneth Carpenter. Evidence of predatory behavior by theropod dinosaurs. „Gaia”. 15, s. 135–144, 1998.
- ↑ Raymond R. Rogers, David W. Krause, Kristina Curry Rogers. Cannibalism in the Madagascan dinosaur Majungatholus atopus. „Nature”. 422 (6931), s. 515–518, 2007. DOI: 10.1038/nature01532. PMID: 12673249.
- ↑ Lars Schmitz, Ryosuke Motani. Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology. „Science”. 332 (6030), s. 705–708, 2011. DOI: 10.1126/science.1200043. PMID: 21493820. Bibcode: 2011Sci...332..705S.
- ↑ Luis M. Chiappe, Lawrence M. Witmer: Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: University of California Press, 2002. ISBN 0-520-20094-2.
- ↑ S. Chatterjee, R.J. Templin. Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui. „Proceedings of the National Academy of Sciences”. 104 (5), s. 1576–1580, 2007. DOI: 10.1073/pnas.0609975104. PMID: 17242354. PMCID: PMC1780066. Bibcode: 2007PNAS..104.1576C.
- ↑ Robert McNeill Alexander. Dinosaur biomechanics. „Proceedings of the Royal Society of Biological Sciences”. 273 (1596), s. 1849–1855, 2006. DOI: 10.1098/rspb.2006.3532. PMID: 16822743. PMCID: PMC1634776.
- ↑ Alain Goriely, Tyler McMillen. Shape of a cracking whip. „Physical Review Letters”. 88 (24), s. 244301, 2002. DOI: 10.1103/PhysRevLett.88.244301. PMID: 12059302. Bibcode: 2002PhRvL..88x4301G.
- ↑ Donald M. Henderson. Effects of stomach stones on the buoyancy and equilibrium of a floating crocodilian: A computational analysis. „Canadian Journal of Zoology”. 81 (8), s. 1346–1357, 2003. DOI: 10.1139/z03-122.
- ↑ a b c d Phil Senter. Voices of the past: a review of Paleozoic and Mesozoic animal sounds. „Historical Biology”. 20 (4), s. 255–287, 2008. DOI: 10.1080/08912960903033327.
- ↑ James A. Hopson. The evolution of cranial display structures in hadrosaurian dinosaurs. „Paleobiology”. 1 (1), s. 21–43, 1975.
- ↑ Carl F. Diegert. A digital acoustic model of the lambeosaurine hadrosaur Parasaurolophus tubicen. „Journal of Vertebrate Paleontology”. 18 (3, Suppl.), s. 38A, 1998.
- ↑ Philip J. Currie, Kevin Padian: Encyclopedia of Dinosaurs. Academic Press, 1997, s. 206.
- ↑ Mark A. Norell i inni, The first dinosaur egg was soft, „Nature”, 2020, DOI: 10.1038/s41586-020-2412-8.
- ↑ Michael H. Hansell: Bird Nests and Construction Behaviour. Cambridge: University of Cambridge Press, 2000. ISBN 0-521-46038-7.
- ↑ David J. Varricchio, John J. Horner, Frankie D. Jackson. Embryos and eggs for the Cretaceous theropod dinosaur Troodon formosus. „Journal of Vertebrate Paleontology”. 22 (3), s. 564–576, 2002. DOI: 10.1671/0272-4634(2002)022[0564:EAEFTC]2.0.CO;2.
- ↑ a b Andrew H. Lee, S Werning. Sexual maturity in growing dinosaurs does not fit reptilian growth models. „Proceedings of the National Academy of Sciences”. 105 (2), s. 582–587, 2008. DOI: 10.1073/pnas.0708903105. PMID: 18195356. PMCID: PMC2206579. Bibcode: 2008PNAS..105..582L.
- ↑ J.R. Horner, Robert Makela. Nest of juveniles provides evidence of family structure among dinosaurs. „Nature”. 282 (5736), s. 296–298, 1979. DOI: 10.1038/282296a0. Bibcode: 1979Natur.282..296H.
- ↑ Luis M. Chiappe, Frankie Jackson, Rodolfo A. Coria: Nesting Titanosaurs from Auca Mahuevo and Adjacent Sites. W: Kristina A. Curry Rogers, Jeffrey A. Wilson: The Sauropods: Evolution and Paleobiology. Berkeley: University of California Press, 2005, s. 285–302. ISBN 0-520-24623-3.
- ↑ Discovering Dinosaur Behavior: 1960–present view [online], Encyclopædia Britannica [zarchiwizowane z adresu 2016-02-22].
- ↑ Qingjin Meng, Jinyuan Liu, David J. Varricchio, Timothy Huang i inni. Parental care in an ornithischian dinosaur. „Nature”. 431 (7005), s. 145–146, 2004. DOI: 10.1038/431145a. PMID: 15356619. Bibcode: 2004Natur.431..145M.
- ↑ Robert R. Reisz, Diane Scott, Hans-Dieter Sues, David C. Evans i inni. Embryos of an Early Jurassic Prosauropod Dinosaur and Their Evolutionary Significance. „Science”. 309 (5735), s. 761–764, 2005. DOI: 10.1126/science.1114942. PMID: 16051793. Bibcode: 2005Sci...309..761R.
- ↑ Neil D.L. Clark, Paul Booth, Cathie Booth, Dugald A. Ross. Dinosaur footprints from the Duntulm Formation (Bathonian, Jurassic) of the Isle of Skye. „Scottish Journal of Geology”. 40 (1), s. 13–21, 2004. DOI: 10.1144/sjg40010013. [dostęp 2011-05-05].
- ↑ Drinking. W: Paul R. Ehrlich, David S. Dobkin, and Darryl Wheye: Birds of Stanford. Standford University, 1988. [dostęp 2007-12-13].
- ↑ Ella Tsahar, Carlos Martínez Del Rio, Ido Izhaki, Zeev Arad. Can birds be ammonotelic? Nitrogen balance and excretion in two frugivores. „Journal of Experimental Biology”. 208 (6), s. 1025–1034, 2005. DOI: 10.1242/jeb.01495. PMID: 15767304.
- ↑ E. Skadhauge, K.H. Erlwanger, S.D. Ruziwa, V. Dantzer i inni. Does the ostrich (Struthio camelus) coprodeum have the electrophysiological properties and microstructure of other birds?. „Comparative biochemistry and physiology. Part A, Molecular & integrative physiology”. 134 (4), s. 749–755, 2003. DOI: 10.1016/S1095-6433(03)00006-0. PMID: 12814783.
- ↑ Marion R. Preest, Carol A. Beuchat. Ammonia excretion by hummingbirds. „Nature”. 386 (6625), s. 561–562, April 1997. DOI: 10.1038/386561a0. Bibcode: 1997Natur.386..561P.
- ↑ J. Mora, J. Martuscelli, J. Ortiz Pineda, G. Soberon. The Regulation of Urea-Biosynthesis Enzymes in Vertebrates. „Biochemical Journal”. 96 (1), s. 28–35, lipiec 1965. PMID: 14343146. PMCID: PMC1206904.
- ↑ Gary C. Packard. The Influence of Ambient Temperature and Aridity on Modes of Reproduction and Excretion of Amniote Vertebrates. „The American Naturalist”. 100 (916), s. 667–682, 1966. DOI: 10.1086/282459. JSTOR: 2459303.
- ↑ Thomas G. Balgooyen. Pellet Regurgitation by Captive Sparrow Hawks (Falco sparverius). „Condor”. 73 (3), s. 382–385, 1971-10-01. DOI: 10.2307/1365774. JSTOR: 1365774.
- ↑ Anusuya Chinsamy, Willem J. Hillenius: Physiology of Nonavian Dinosaurs. W: David B. Weishampel, Peter Dodson, Halszka Osmólska: The Dinosauria (2nd edition). University of California Press, 2004, s. 643–659. ISBN 0-520-24209-2.
- ↑ John R. Hutchinson: Hot-Blooded or Cold-Blooded??. University of California Museum of Paleontology, 1995-11-28. [dostęp 2012-02-12]. (ang.).
- ↑ Keith M. Parsons: Drawing Out Leviathan: Dinosaurs and the Science Wars. Bloomington: Indiana University Press, 2001, s. 22–48. ISBN 0-253-33937-5.
- ↑ a b c Paul C. Sereno, Ricardo N. Martinez, Jeffrey A. Wilson, David J. Varricchio i inni. Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina. „PLoS ONE”. 3 (9), s. e3303, 09 2008. DOI: 10.1371/journal.pone.0003303. PMID: 18825273. PMCID: PMC2553519. Bibcode: 2008PLoSO...3.3303S.
- ↑ R.E.H. Reid: Dinosaurian Physiology: the Case for „Intermediate” Dinosaurs. W: James O. Farlow, M.K. Brett-Surman: The Complete Dinosaur. Bloomington: Indiana University Press, 1997, s. 449–473. ISBN 0-253-33349-0.
- ↑ Maria E. McNamara i inni, Fossilized skin reveals coevolution with feathers and metabolism in feathered dinosaurs and early birds, „Nature Communications”, 9 (1), 2018, DOI: 10.1038/s41467-018-04443-x, ISSN 2041-1723, PMID: 29802246, PMCID: PMC5970262 [dostęp 2024-06-12] (ang.).
- ↑ Thomas H. Huxley. On the animals which are most nearly intermediate between birds and reptiles. „Annals of the Magazine of Natural History”. 4 (2), s. 66–75, 1868.
- ↑ Gerhard Heilmann: The Origin of Birds. London: Witherby, 1926, s. 208. ISBN 0-486-22784-7.
- ↑ Henry Fairfield Osborn. Three new Theropoda, Protoceratops zone, central Mongolia. „American Museum Novitates”. 144, s. 1–12, 1924.
- ↑ John H. Ostrom. The ancestry of birds. „Nature”. 242 (5393), s. 136, 1973. DOI: 10.1038/242136a0. Bibcode: 1973Natur.242..136O.
- ↑ Jacques Gauthier: Saurischian Monophyly and the Origin of Birds. W: Kevin Padian: The Origin of Birds and the Evolution of Flight. 1986, s. 1–55, seria: Memoirs of the California Academy of Sciences 8.
- ↑ Gerald Mayr, Burkhard Pohl, Stefan Peters. A Well-Preserved Archaeopteryx Specimen with Theropod Features. „Science”. 310 (5753), s. 1483–1486, 2005. DOI: 10.1126/science.1120331. PMID: 16322455. Bibcode: 2005Sci...310.1483M.
- ↑ Larry D. Martin. A basal archosaurian origin for birds. „Acta Zoologica Sinica”. 50 (6), s. 977–990, 2006.
- ↑ Peter Wellnhofer. Ein neuer Exemplar von Archaeopteryx. „Archaeopteryx”. 6, s. 1–30, 1988.
- ↑ Phil Senter. A new look at the phylogeny of Coelurosauria (Dlnosauria: Theropoda). „Journal of Systematic Palaeontology”. 5 (4), s. 429–463, 2007. DOI: 10.1017/S1477201907002143. (ang.).
- ↑ Xing Xu, Mark A. Norell, Xuewen Kuang, Xiaolin Wang i inni. Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids. „Nature”. 431 (7009), s. 680–684, 200 4. DOI: 10.1038/nature02855. PMID: 15470426. Bibcode: 2004Natur.431..680X.
- ↑ Ursula B. Göhlich, Luis M. Chiappe. A new carnivorous dinosaur from the Late Jurassic Solnhofen archipelago. „Nature”. 440 (7082), s. 329–332, 2006. DOI: 10.1038/nature04579. PMID: 16541071. Bibcode: 2006Natur.440..329G.
- ↑ Theagarten Lingham-Soliar. The dinosaurian origin of feathers: perspectives from dolphin (Cetacea) collagen fibers. „Naturwissenschaften”. 90 (12), s. 563–567, 2003. DOI: 10.1007/s00114-003-0483-7. PMID: 14676953. Bibcode: 2003NW.....90..563L.
- ↑ a b Alan Feduccia, Theagarten Lingham-Soliar, J. Richard Hinchliffe. Do feathered dinosaurs exist? Testing the hypothesis on neontological and paleontological evidence. „Journal of Morphology”. 266 (2), s. 125–166, 2005. DOI: 10.1002/jmor.10382. PMID: 16217748.
- ↑ Theagarten Lingham-Soliar, Alan Feduccia, Xiaolin Wang. A new Chinese specimen indicates that ‘protofeathers’ in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres. „Proceedings of the Biological Sciences”. 274 (1620), s. 1823–1829, 2007. DOI: 10.1098/rspb.2007.0352. PMID: 17521978. PMCID: PMC2270928.
- ↑ Richard O. Prum. Are Current Critiques Of The Theropod Origin Of Birds Science? Rebuttal To Feduccia 2002. „The Auk”. 120 (2), s. 550–561, April 2003. DOI: 10.1642/0004-8038(2003)120[0550:ACCOTT]2.0.CO;2. JSTOR: 4090212.
- ↑ Patrick M. O’Connor, Leon P.A.M. Claessens. Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs. „Nature”. 436 (7048), s. 253–256, 2005. DOI: 10.1038/nature03716. PMID: 16015329. Bibcode: 2005Natur.436..253O.
- ↑ Meat-Eating Dinosaur from Argentina Had Bird-Like Breathing System. [dostęp 2011-05-05].
- ↑ Xing Xu, Mark A. Norell. A new troodontid dinosaur from China with avian-like sleeping posture. „Nature”. 431 (7010), s. 838–841, 2004. DOI: 10.1038/nature02898. PMID: 15483610. Bibcode: 2004Natur.431..838X.
- ↑ Mark A. Norell, James M. Clark, Luis M. Chiappe, Demberelyln Dashzeveg. A nesting dinosaur. „Nature”. 378 (6559), s. 774–776, 1995. DOI: 10.1038/378774a0. Bibcode: 1995Natur.378..774N.
- ↑ David J. Varricchio, Jason R. Moore, Gregory M. Erickson, Mark A. Norell i inni. Avian Paternal Care Had Dinosaur Origin. „Science”. 322 (5909), s. 1826–1828, 2008. DOI: 10.1126/science.1163245. PMID: 19095938. Bibcode: 2008Sci...322.1826V.
- ↑ Oliver Wings. A review of gastrolith function with implications for fossil vertebrates and a revised classification. „Palaeontologica Polonica”. 52 (1), s. 1–16, 2007. [dostęp 2011-05-05].
- ↑ Lowell Dingus, Timothy Rowe: The Mistaken Extinction – Dinosaur Evolution and the Origin of Birds. New York: W.H. Freeman, 1998. ISBN 978-0-7167-2944-0.
- ↑ Kenneth G. Miller, Michelle A. Kominz, James V. Browning, James D. Wright i inni. The Phanerozoic Record of Global Sea-Level Change. „Science”. 310 (5752), s. 1293–1298, 2005. DOI: 10.1126/science.1116412. PMID: 16311326. Bibcode: 2005Sci...310.1293M.
- ↑ J.M. McArthur, N.M.M. Janssen, S. Reboulet, M.J. Leng i inni. Palaeotemperatures, polar ice-volume, and isotope stratigraphy (Mg/Ca, δ18O, δ13C, 87Sr/86Sr): The Early Cretaceous (Berriasian, Valanginian, Hauterivian). „Palaeogeography, Palaeoclimatology, Palaeoecology”. 248 (3–4), s. 391–430, 2007. DOI: 10.1016/j.palaeo.2006.12.015.
- ↑ Gregory S. Paul: Dinosaurs of the air: the evolution and loss of flight in dinosaurs and birds. Johns Hopkins University Press, 2002, s. 397. ISBN 0-8018-6763-0.
- ↑ Luis W. Alvarez, Walter Alvarez, Frank Asaro, Helen V. Michel. Extraterrestrial cause for the Cretaceous–Tertiary extinction. „Science”. 208 (4448), s. 1095–1108, 1980. DOI: 10.1126/science.208.4448.1095. PMID: 17783054. Bibcode: 1980Sci...208.1095A.
- ↑ Alan R. Hildebrand, Glen T. Penfield, David A. Kring, Mark Pilkington i inni. Chicxulub Crater; a possible Cretaceous/Tertiary boundary impact crater on the Yucatan Peninsula, Mexico. „Geology”. 19 (9), s. 867–871, September 1991. DOI: 10.1130/0091-7613(1991)019<0867:CCAPCT>2.3.CO;2. Bibcode: 1991Geo....19..867H.
- ↑ Kevin O. Pope, Adriana C. Ocampo, Gary L. Kinsland, Randy Smith. Surface expression of the Chicxulub crater. „Geology”. 24 (6), s. 527–530, 1996. DOI: 10.1130/0091-7613(1996)024<0527:SEOTCC>2.3.CO;2. PMID: 11539331. Bibcode: 1996Geo....24..527P.
- ↑ D.S. Robertson, et al. Survival in the first hours of the Cenozoic. „Geological Society of America Bulletin”. 116 (5/6), s. 760–768, 2003. DOI: 10.1130/B25402.1.
- ↑ Philippe Claeys, Steven Goderis. Solar System: Lethal billiards. „Nature”. 449 (7158), s. 30–31, 2007-09-05. DOI: 10.1038/449030a. PMID: 17805281. Bibcode: 2007Natur.449...30C.
- ↑ Tammy Plotner: Did Asteroid Baptistina Kill the Dinosaurs? Think other WISE .... [w:] Universe Today [on-line]. 2011. [dostęp 2011-09-20].
- ↑ Christian Koeberl, Kenneth G. MacLeod: Catastrophic Events and Mass Extinctions. Geological Society of America, 2002. ISBN 0-8137-2356-6. OCLC 213836505.
- ↑ a b C. Hofmann, G. Féraud, V. Courtillot. 40Ar/39Ar dating of mineral separates and whole rocks from the Western Ghats lava pile: further constraints on duration and age of the Deccan traps. „Earth and Planetary Science Letters”. 180, s. 13–27, 2000. DOI: 10.1016/S0012-821X(00)00159-X. Bibcode: 2000E&PSL.180...13H.
- ↑ a b c R.A. Duncan, D.G. Pyle. Rapid eruption of the Deccan flood basalts at the Cretaceous/Tertiary boundary. „Nature”. 333 (6176), s. 841–843, 1988. DOI: 10.1038/333841a0. Bibcode: 1988Natur.333..841D.
- ↑ Walter Alvarez: T. rex and the Crater of Doom. Princeton University Press, 1997, s. 130–146. ISBN 978-0-691-01630-6.
- ↑ J.E. Fassett, S.G. Lucas, R.A. Zielinski, J.R. Budahn. Compelling new evidence for Paleocene dinosaurs in the Ojo Alamo Sandstone, San Juan Basin, New Mexico and Colorado, USA. „Catastrophic events and mass extinctions, Lunar and Planetary Contribution”. 1053, s. 45–46, 2001. Bibcode: 2001caev.conf.3139F. [dostęp 2007-05-18].
- ↑ Robert E. Sloan, J. Keith Rigby Jr., Leigh M. Van Valen, Diane Gabriel. Gradual dinosaur extinction and simultaneous ungulate radiation in the Hell Creek Formation. „Science”. 232 (4750), s. 629–633, 1986. DOI: 10.1126/science.232.4750.629. PMID: 17781415. Bibcode: 1986Sci...232..629S.
- ↑ a b David E. Fastovsky, Peter M. Sheehan. Reply to comment on „The Extinction of the dinosaurs in North America”. „GSA Today”. 15 (7), s. 11, 2005. DOI: 10.1130/1052-5173(2005)015[11b:RTEOTD]2.0.CO;2.
- ↑ Robert M. Sullivan. No Paleocene dinosaurs in the San Juan Basin, New Mexico. „Geological Society of America Abstracts with Programs”. 35 (5), s. 15, 2003. [dostęp 2007-07-02]. [zarchiwizowane z adresu].
- ↑ James E. Fassett, Larry M. Heaman, Antonio Simonetti. Direct U–Pb dating of Cretaceous and Paleocene dinosaur bones, San Juan Basin, New Mexico. „Geology”. 39 (2), s. 159–162, 2011. DOI: 10.1130/G31466.1.
- ↑ Dong Zhiming: Dinosaurian Faunas of China. China Ocean Press, Beijing, 1992. ISBN 3-540-52084-8. OCLC 26522845.
- ↑ Dinosaur bones ‘used as medicine’. BBC News, 2007-07-06. [dostęp 2007-07-06].
- ↑ a b William A.S. Sarjeant: The Earliest Discoveries. W: M.K. Brett-Surman, Thomas R. Holtz Jr, James O. Farlow: The Complete Dinosaur. Bloomington: Indiana University Press, 1997, s. 3–11. ISBN 0-253-33349-0.
- ↑ Edward Lhuyd: Lithophylacii Britannici Ichnographia, sive lapidium aliorumque fossilium Britannicorum singulari figura insignium. London: Gleditsch and Weidmann, 1699.
- ↑ Justin B. Delair, William A.S. Sarjeant. The earliest discoveries of dinosaurs: the records re-examined. „Proceedings of the Geologists’ Association”. 113 (3), s. 185–197, 2002. DOI: 10.1016/S0016-7878(02)80022-0.
- ↑ R.T. Gunther: Life and letters of Edward Lhwyd: Second keeper of the Museum Ashmoleanum (Early science in Oxford Volume XIV). Dawsons of Pall Mall, 1968.
- ↑ William Buckland. Notice on the Megalosaurus or great Fossil Lizard of Stonesfield. „Transactions of the Geological Society of London”. 1 (2), s. 390–396, 1824. DOI: 10.1144/transgslb.1.2.390.
- ↑ Gideon A. Mantell. Notice on the Iguanodon, a newly discovered fossil reptile, from the sandstone of Tilgate forest, in Sussex. „Philosophical Transactions of the Royal Society”. 115, s. 179–186, 1825. DOI: 10.1098/rstl.1825.0010. JSTOR: 107739.
- ↑ Hans-Dieter Sues: European Dinosaur Hunters. W: James O. Farlow, M.K. Brett-Surman: The Complete Dinosaur. Bloomington: Indiana University Press, 1997, s. 14. ISBN 0-253-33349-0.
- ↑ Nicolaas A. Rupke: Richard Owen: Victorian Naturalist. New Haven: Yale University Press, 1994. ISBN 978-0-300-05820-8.
- ↑ Thom Holmes: Fossil Feud: The Bone Wars of Cope and Marsh, Pioneers in Dinosaur Science. Silver Burdett Press, 1996. ISBN 978-0-382-39147-7. OCLC 34472600.
- ↑ Robert T. Bakker: The Dinosaur Heresies. New York: William Morrow, 1986. ISBN 0-8217-5608-7.
- ↑ Nick Evershed, Blood, tissue extracted from duck-billed dinosaur bone [online], Cosmos Online, 1 maja 2009 [zarchiwizowane z adresu 2013-10-05] (ang.).
- ↑ Hai-Lin Wang, Zi-Ying Yan, Dong-Yan Jin. Reanalysis of published DNA sequence amplified from Cretaceous dinosaur egg fossil. „Molecular Biology and Evolution”. 14 (5), s. 589–591, 1997-05-01. DOI: 10.1093/oxfordjournals.molbev.a025796. PMID: 9159936. [dostęp 2007-12-05].
- ↑ Belinda S.W. Chang, Karolina Jönsson, Manija A. Kazmi, Michael J. Donoghue i inni. Recreating a Functional Ancestral Archosaur Visual Pigment. „Molecular Biology and Evolution”. 19 (9), s. 1483–1489, 2002-09-01. DOI: 10.1093/oxfordjournals.molbev.a004211. PMID: 12200476. [dostęp 2007-12-05].
- ↑ Mary H. Schweitzer, Mark Marshall, Keith Carron, D. Scott Bohle i inni. Heme compounds in dinosaur trabecular bone. „Proceedings of the National Academy of Sciences”. 94 (12), s. 6291–6296, 1997. DOI: 10.1073/pnas.94.12.6291. PMID: 9177210. PMCID: PMC21042. Bibcode: 1997PNAS...94.6291S.
- ↑ Graham Embery, Angela C. Milner, Rachel Waddington, Rachel C. Hall i inni. Identification of proteinaceous material in the bone of the dinosaur Iguanodon. „Connect Tissue Res”. 44 (Suppl 1), s. 41–46, 2003. DOI: 10.1080/713713598. PMID: 12952172.
- ↑ Joseph E. Peterson, Melissa E. Lenczewski, Reed P. Scherer. Influence of Microbial Biofilms on the Preservation of Primary Soft Tissue in Fossil and Extant Archosaurs. „PLoS ONE”. 5 (10), s. 13A, October 2010. DOI: 10.1371/journal.pone.0013334.
- ↑ Dinosaur – Definition and More. Merriam-Webster Dictionary. [dostęp 2011-05-06].
- ↑ H.S. Torrens. The dinosaurs and dinomania over 150 years. „Modern Geology”. 18 (2), s. 257–286, 1993.
- ↑ Brent H. Breithaupt: First golden period in the USA. W: Philip J. Currie, Kevin Padian: The Encyclopedia of Dinosaurs. San Diego: Academic Press, 1997, s. 347–350. ISBN 978-0-12-226810-6.
- ↑ „London. Michaelmas term lately over, and the Lord Chancellor sitting in Lincoln’s Inn Hall. Implacable November weather. As much mud in the streets, as if the waters had but newly retired from the face of the earth, and it would not be wonderful to meet a Megalosaurus, forty feet long or so, waddling like an elephantine lizard up Holborne Hill.” Dickens CJH: Bleak House. London: Bradbury & Evans, 1852, s. 1.
- ↑ Donald F. Glut, M.K. Brett-Surman: Dinosaurs and the Media. W: James O. Farlow, M.K. Brett-Surman: The Complete Dinosaur. Indiana University Press, 1997, s. 675–697. ISBN 978-0-253-21313-6.
Bibliografia
edytuj- Richard Owen, Report on British Fossil Reptiles. Part II. Report of the Eleventh Meeting of the British Association for the Advancement of Science; Held at Plymouth in July 1841, London: John Murray, 1842, s. 60–204.
Linki zewnętrzne
edytuj- Karolina Głowacka i Daniel Tyborowski, Czy dinozaury stworzyłyby kiedyś cywilizację?, kanał „Radio Naukowe” na YouTube, 25 marca 2021 [dostęp 2023-09-06].