爬虫類の一グループとして分類され、三畳紀中期(2億3500万年以上前)から白亜紀末(6600万年前)まで繁栄した生物。ハトほどの大きさから全長30m以上まで様々な大きさ・形態のものが現れ、鳥類を残して絶滅した。便宜上、恐竜は爬虫類とされているが、トカゲやワニではなく鳥類が恐竜の生き残りである。逆にいえば、鳥類は呼び方が変わっただけで恐竜そのもの、つまり爬虫類であるとも言える。爬虫類の定義が元々曖昧なのである。
定義と特徴
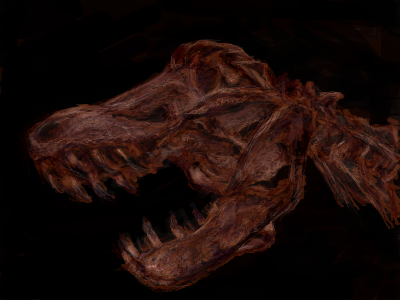
|
| ティラノサウルス頭骨 |
日常的な会話では、分類学上の恐竜に限らず翼竜・魚竜・首長竜、場合によってはマンモスやイクチオステガまで、絶滅して現在似たものがいない爬虫類または四肢動物のこと全般が「恐竜」と呼ばれがちである。
しかし生物学上ではより狭い範囲に限定されており、定義の表現し方は二通りある。
- 解剖学的な定義:後肢が前から見て地面に垂直になるような骨盤の構造をしており、その骨盤に3つ以上の脊椎のパーツ(仙椎)がある爬虫類が恐竜である。
- 分岐学的な定義:鳥とトリケラトプスそれぞれから祖先をさかのぼっていき、最初に一致した動物から進化した生物は全て恐竜である。
解剖学的な定義の方が先にあったものだが、解剖学的な定義は分岐学的な定義を具体的にどんな動物を指すのか言い換えたものといえる。分岐学的な定義では鳥類はすでに恐竜に含まれている。単に恐竜と言った場合、「非鳥類恐竜」を指すことが多い。
また恐竜といえば巨大なものと思いがちだが、実際は大きさは関係ない。鳥類を含めて恐竜と言った場合、最小の恐竜はマメハチドリ(5cm、2g)である。鳥類を除くと、数十cmほどのものが何種類か見つかっている(動物全体から見るとそれなりに大きいとは言える)。
羽毛が生えている恐竜がいることや恐竜から進化したということは1980年代から言われてきたことではあるが、もはや仮説ではなく羽毛が生えている恐竜がいること含め、学会でも常識である。
現在の爬虫類の中ではワニが近縁で、翼竜などいくつかの他の絶滅したものと合わせて主竜類というグループを成す。
元々爬虫類は後肢がほぼ左右に向かって伸び、這いつくばった姿勢であった。しかし主竜類は後肢が地面に垂直になる(直立歩行という)傾向があった。主竜類の中から二足歩行する小型のものが現れ、さらに完全に直立歩行になることで恐竜に進化した。
四足歩行の恐竜も二足歩行の名残りで重心が後肢に寄っており、前肢が短いものが多かった。また哺乳類に比べると胴体の柔軟性が低く、筋肉より腱や骨格で胴体を支える傾向があった。
二足歩行だった、または四足歩行でも重心が腰に寄っていたのは、直立歩行になったことで後肢を後ろに蹴ったときに胴体を起こすような反動が発生することに対応したためと考えられる。後肢を蹴る反動により前肢が浮きやすくなったため、曲がりにくい胴体を水平に保ちながら後肢だけで歩くようになったようだ。哺乳類の場合は逆に前肢が浮かないように重量を分配していて、チーターのようにしなやかな胴体の動きが四肢と連動しているものまでいる。
直立歩行により体重を支えやすく、また長時間移動しやすくなったことは他の這いつくばった姿勢の爬虫類に対するアドバンテージとなった。しかし三畳紀後期の環境変化を乗り切って繁栄することができたのは、排泄に要する水分の節約、高い呼吸能力など、他の爬虫類にない当時の環境に適合した特徴のためであると言われる。
歴史
三畳紀中期~後期(2億5200万年前~)
全ての大陸が地続きになって「超大陸パンゲア」を形成、陸上生物は分布を広げやすかった。植物はシダ植物や裸子植物(針葉樹やイチョウ等の仲間)が中心だった。現代のような草原はずっと後まで現れない。
単弓類や、主竜類などの様々な爬虫類が繁栄しており、単弓類からは哺乳類、主竜類からはワニ、翼竜、そして恐竜が進化した。
特に原始的な恐竜は主にブラジルで見つかっており、恐竜の起源は南米にあったと考えられている。しかし足跡化石によると、三畳紀前期に他の地域で誕生していた可能性もある。最初の恐竜は小型で二足歩行だったと考えられる。
捕食者としてライバルの多い獣脚類はおおむね4m以下と小型だったが、一部は6mほどに達した。竜脚形類に属する一部の植物食恐竜はすでに10mほどまで大型化していた。
ジュラ紀前期~中期(2億100万年前~)
三畳紀末の大絶滅により、他の陸生爬虫類は減少。ケラトサウルス類や原始的なテタヌラ類に属する肉食恐竜が、本格的に大型化を始めた。この頃にはすでにコエルロサウルス類に属する鳥類に近縁な恐竜がいたという間接的な証拠がある。
プラテオサウルス類などの竜脚形類の植物食恐竜が繁栄したが、その中で原始的な竜脚類も多様化しつつあった。
ジュラ紀後期(1億6350万年前~)
超大陸パンゲアが、南のゴンドワナと北のローラシアに分離しつつあった。高二酸化炭素濃度・低湿度という環境下で針葉樹は大きく生長したが栄養価は低くなった。
マメンチサウルスやアパトサウルスなど、ジュラ紀後期特有の竜脚類が本格的に繁栄・大型化し、一部は全長30mを超えた。また剣竜類や鳥脚類も繁栄し、一部は大型化。それに合わせるように、ケラトサウルス類・メガロサウルス類・アロサウルス類に属する、典型的な大型肉食恐竜も繁栄した。
また、小型の恐竜の多様性も増し、原始的なティラノサウルス類や鳥類を含んだ羽毛恐竜(コエルロサウルス類)に代表される、後の白亜紀に繁栄することになる恐竜も現れ始めていた。
白亜紀前期(1億4500万年前~)
大陸の分裂が進み、これ以降陸生動物の分布は、その進化の歴史上で大陸のつながりがどうなっていたかに大きく左右されることになった。温暖な気候の元海面がかなり上昇し、浅い海が広がった。またアルカエフルクトゥスのような被子植物(目立つ花の咲く植物)も見られるようになった。
ジュラ紀的な竜脚類や剣竜類はほとんど姿を消し、マクロナリアに属する白亜紀的な竜脚類や鳥脚類が繁栄した。また、鎧竜類や原始的な角竜類も多様化しつつあった。アロサウルス類の肉食恐竜は引き続き繁栄していた。ケラトサウルス類と、メガロサウルス類から進化したスピノサウルス類は、南のゴンドワナ由来の大陸で特殊化し独自の地位に着いた。
鳥類やコエルロサウルス類はさらに多様性を増した。その中には、すでに高度な飛行能力を得た鳥、肉食を捨て植物食に適応したものまでいた。
白亜紀後期(1億50万年前~)
インド亜大陸が未だアジアと分離していたりする以外は、ほぼ現在の大陸の分割が確立した。引き続き温暖な気候で被子植物が繁栄し、これは植物食恐竜の進化にも影響を与えたと考えられる。
アジア・北米では、鳥脚類・鎧竜類・堅頭竜類・角竜類が大繁栄した。アロサウルス類は姿を消し、ティラノサウルス類が大型化して主要な捕食者の位置についた。コエルロサウルス類は繁栄を極め、現在と変わらない鳥類も現れた。
アフリカや南米では、北半球的な恐竜の一部が進入していたが、引き続きマクロナリアの竜脚類と、ケラトサウルス類・スピノサウルス類・アロサウルス類の獣脚類が繁栄し、一部はかなり大型化した。マクロナリアはアジアや北米にも若干見られた。
新生代(6600万年前~)
一部の鳥類を除く全ての恐竜が絶滅(「絶滅について」を参照)。鳥類だけはその後も繁栄を続けた。
各分類群の特徴
骨盤の特徴によって竜盤類と鳥盤類に大別され、さらに以下のように分かれる。この系統樹に例示した個別の恐竜には個別の記事があり、それぞれの分類群について詳述している。
恐竜┬竜盤類┬獣脚類:アロサウルス、ティラノサウルス、ヴェロキラプトル、始祖鳥など
│ └竜脚形類:アパトサウルスなど
│
└鳥盤類┬装盾類┬剣竜類:ステゴサウルスなど
│ └鎧竜類:アンキロサウルスなど
│
└角脚類┬周飾頭類┬堅頭竜類:パキケファロサウルスなど
│ └角竜類:トリケラトプスなど
│
└鳥脚類:パラサウロロフスなど
鳥盤類は全てクチバシを持ち骨盤のシルエットが鳥のものに似ていたが、鳥類を輩出したのは竜盤類に含まれる獣脚類である。
「代表的な種類」には、それぞれの分類群のさらに細かい分類とその中でも重要な種類(有名なもの、国内産のもの、なるべく各グループの進化を辿れるもの、重要な情報をもたらしたもの、お絵カキコのあるもの)、その分類群の特徴を挙げた。分類は研究者の意見によって異なることもあり、またここで使っている分類は必ずしも正確ではなく、細かいグループを全て網羅しているものでもない。
名前の後の■は記事に直接使っていないお絵カキコへのリンク(◆は種類を特定していないお絵カキコ)。
獣脚類

|
| アロサウルス |
いわゆる「肉食恐竜」と言われるもの。
恐竜の代表格ティラノサウルスから鳥類にごく近縁なもの、あるいは鳥類自体まで非常に様々な種類を含む。
二足歩行。前後に長い頭部、長めの首、カプセル状の胴体、長い後肢と尾、短めの前肢、鋭い牙、または歯のないクチバシを持つ。羽毛を持っていたとされるものも多い。羽毛以外にも骨格に鳥類的な特徴が多い。また恐竜の中では体重の割に特に脳容積が大きく、一部のものでは鳥類の水準に達していた。
一般に大型のものは腕よりも顎の方が発達しており、頭部が大型化していても穴が多く軽量で、衝撃を吸収する構造になっていた。
大型種が狩りをした場合は、腕で獲物を押さえつけるよりも顎で肉を大きく削ぎ取ったか噛み砕いたと考えられている。
またクチバシがあり歯がないもの、歯があっても肉を切り裂けないような形のものは二次的に植物食に適応したのだと言われる。
代表的な種類
| ヘレラサウルス類 | ヘレラサウルス■、フレングエリサウルス、スタウリコサウルス | 特に原始的。獣脚類に含めないことが多い。 |
| 基盤的獣脚類/コエロフィシス類 | エオドロマエウス/コエロフィシス、リリエンステルヌス、ポドケサウルス、メガプノサウルス、ディロフォサウルス | 細長い体形をしており、小型から中型。 |
| ケラトサウルス類 | ケラトサウルス■、エラフロサウルス、カルノタウルス■、マジュンガサウルス、マシアカサウルス■ | ノアサウルス科を除きやや大型。手指が4本残っている。 |
| メガロサウルス類 | ピアトニツキサウルス、エウストレプトスポンディルス、メガロサウルス、トルヴォサウルス、アフロヴェナトル | ここから下は尾があまり曲がらないテタヌラ類。やや後肢の短いオーソドックスな肉食恐竜。 |
| スピノサウルス類 | バリオニクス、スコミムス、イリタトル、スピノサウルス■ | 長い吻部と円錐形の歯を持ち、魚食に適応したとみられる。メガロサウルス類を含めることが多い。 |
| アロサウルス類 | モノロフォサウルス■、アロサウルス■、ヤンチュアノサウルス、フクイラプトル、コンカヴェナトル■、アクロカントサウルス、カルカロドントサウルス | 標準的な中型~大型肉食恐竜。やや軽量な体つきで、歯が鋭く前肢親指の爪が大きい。 |
| コンプソグナトゥス類 | コンプソグナトゥス、ジュラヴェナトル、スキピオニクス、シノサウロプテリクス■、シノカリオプテリクス■ | ここから下がコエルロサウルス類。小型で身軽。原羽毛を持つ。 |
| ティラノサウルス類 | グアンロング■、ラプトレックス■、アルバートサウルス、テラトフォネウス■、タルボサウルス、ナノティランヌス■、ティラノサウルス■■ | 最初は他のコエルロサウルス類と大差なかったが次第に独自に大型化した。前肢が特に小さい。大型のものほどがっしりとした体形。 |
| オルニトミムス類 | ペレカニミムス、デイノケイルス、ガルディミムス、ガリミムス、オルニトミムス | いわゆるダチョウ恐竜。クチバシを持つ。後肢や首だけでなく手も長い。雑食及び葉食、またはプランクトン食とされる。 |
| オヴィラプトル類 | インキシヴォサウルス、カウディプテリクス、アジャンシンゲニア、オヴィラプトル◆■、キロステノテス | 丸っこい頭と高さのあるクチバシ、短い尾、大きな手を持つ。雑食及び果実食とされる。 |
| テリジノサウルス類 | ファルカリウス、セグノサウルス、アラシャサウルス、ノスロニクス、テリジノサウルス■■ | どっしりした胴体、巨大な鉤爪など特異な体形。植物食とされる。 |
| アルヴァレスサウルス類 | ハプロケイルス、アルヴァレスサウルス、パタゴニクス、モノニクス、シュヴウイア | オルニトミムス類に似るがずっと小型で前肢はとても太短く、手指は大きな爪のある1本以外退化。 |
| エウマニラプトラ | エピデンドロサウルス◆、アンキオルニス■、トロオドン■、ヴェロキラプトル◆、デイノニクス、ミクロラプトル、始祖鳥■、ハシビロコウさん | いわゆるラプトル。原始的な鳥類であるともされるほど鳥に近い。分岐学上は鳥自体も含む。 |
竜脚形類

|
| アパトサウルス |
「恐竜」のイメージどおりの姿をしたもののグループ。ジュラ紀中期から白亜紀にかけて巨大化し、一部のものは全長30mを超えたことは確実とされる。
特に原始的なものの一部を除き四足歩行、植物食性。小さな頭、長い首と尾、樽状の胴体、柱のような四肢を持つ。
骨格には大きな体を軽量化しつつも少ない力で確実に支える適応が多く見られる。頭部は長い首の先にあるだけに軽量で咀嚼力は弱い。
筋肉の力より骨自体の構造や腱の引っ張りで体勢を保っていた。長い首でキリンのように高い枝の葉を食べたと言われてきたが、実際には長い首はあまり上に向かず、つり橋のように背中側から靭帯で支えて前に伸ばし、ゾウの鼻のように広い範囲の植物を食べることに役立てていた可能性が高い。
丸呑みした植物を長い腸でゆっくりと消化していたとされる。
狭義の竜脚類に含まれない初期のものはこの限りではなく、二足歩行など独自路線を辿っていた。
代表的な種類
| ごく原始的なもの | エオラプトル、パンファギア、グアイバサウルス、サトゥルナリア | 肉食恐竜と大差ない、小型で二足歩行のランナー。 |
| プラテオサウルス類など | テコドントサウルス、プラテオサウルス、リオハサウルス、マッソスポンディルス、アンキサウルス | 二足または四足歩行。独自の路線で植物食に適応した。古竜脚類と呼ばれ後の竜脚類の祖先ではないとされてきた。 |
| 原始的竜脚類 | アンテトニトルス、イサノサウルス、ヴルカノドン、シュノサウルス、ケティオサウルス | 四足歩行に適応したが体を支える仕組みなどは未発達。 |
| マメンチサウルス類 | ベルサウルス、チュアンジエサウルス、クラメリサウルス、マメンチサウルス、オメイサウルス | アジアに特有。首が長く、体長の半分に達するものも。 |
| ディプロドクス類 | ディプロドクス、アパトサウルス、スーパーサウルス、ディクラエオサウルス、アマルガサウルス■、ニジェールサウルス■ | 細長い尾、長い顔面、やや短い前肢、高い背中を持つ。 |
| マクロナリア | カマラサウルス、ブラキオサウルス■、タンバティタニス、フクイティタン、ティタノサウルス、アルゼンチノサウルス、アラモサウルス | 従来竜脚類のあまり見つかっていなかった白亜紀に特に繁栄。バランスのとれた体形だが装甲のあるものなども。 |
剣竜類

|
| ステゴサウルス |
植物食、四足歩行。クチバシのある小さな頭、縦に大きな胴体、比較的長い尾と後肢・短い前肢、背や尾に並ぶ骨の棘や板を持つ。
棘や骨板の表面に溝が多数見つかっており、背中にしっかり固定するためのコラーゲン繊維や板表面に血液をめぐらすための血管の痕跡と見られる。鳥盤類としては歯や顎が貧弱だった。
棘や骨板の役割には諸説あるが、棘は防御や外敵の撃退、板は放熱または日光からの受熱、同種へのディスプレイに用いられたと思われる。
地表面の柔らかい植物を食べたと考えられる。
肉食恐竜アロサウルスの化石のなかには、ステゴサウルスの棘が見事にはまるケガのあるものが複数見つかっており、アロサウルスとステゴサウルスの間の闘争を示す証拠とされている。
代表的な種類
| 基盤的装盾類 | スクテロサウルス■、スケリドサウルス | やや身軽。装甲が背中全体に散らばり鎧竜類にも似ていて、剣竜類と鎧竜類両方の祖先とされる。 |
| ファヤンゴサウルス類 | ファヤンゴサウルス | 原始的で、ステゴサウルス類と違ってやや横幅のある体形。 |
| ステゴサウルス類 | ダケントルルス、ミラガイア、トゥオジャンゴサウルス、ケントロサウルス、ステゴサウルス | 背中の中央に骨板や棘が発達。 |
鎧竜類

|
| アンキロサウルス |
植物食、四足歩行、鎧や棘に覆われた幅の広い胴体と頭、短い四肢を持つ。
口先や鼻先の形態は意外と多様で、鼻の奥が哺乳類のように発達した種類もある。歯や咬筋は発達していない。
ノドサウルス類とアンキロサウルス類に二分されてきたが、ポラカントゥス類を独立させる意見もある。
アンキロサウルス類の腱で硬化した尾の先にはハンマーがあり、またハンマーのないノドサウルス類でも、大きな棘を肩に発達させたものが多かった。
ハンマーは成体のものなら充分肉食恐竜を撃退する威力があったと言われる。またノドサウルス類も肩の棘を向けて脅すことで外敵を追い払えただろう。
種類によって異なった植物を食べたかもしれない。鼻は呼吸気の温度や湿度を調節したり鳴き声を上げるのに使ったと思われる。
扁平な胴体や顔面の形態から、カバのような半水生動物として復元される場合もある。
代表的な種類
| ノドサウルス類 | アニマンタルクス、サウロペルタ、ノドサウルス、エドモントニア | ハンマーはない。装甲にむらがなく、軽量化されておらず重装甲。 |
| ポラカントゥス類 | ガーゴイロサウルス、ポラカントゥス、ガストニア■、ミモオラペルタ | 前後に短い頭骨からアンキロサウルス類に近縁とされるがハンマーはない。装甲がやや腰に集中する傾向があった。 |
| アンキロサウルス類 | ミンミ、アンキロサウルス、エウオプロケファルス、ピナコサウルス、サイカニア | ハンマー以外にも特に幅広い胴体が特徴。装甲は裏が肉抜きされている。 |
堅頭竜類

|
| ホマロケファレ |

|
| パキケファロサウルス |
丸い頭で親しまれる、いわゆる「石頭恐竜」。
植物食、二足歩行。後頭部の発達した頑丈な頭、幅の広い胴体、太めの尾、長い後肢と短い前肢を持つ。
頭部は種類または個体によって様々な形の角やドームで飾られている。ドームの内部構造も頭突きで闘争する現在の植物食動物に似ていた。頸椎は細かったが首の肋骨(頸肋骨)は太長く発達していた。
代表的な種類として挙げたもののうちスティギモロクとドラコレックスはそれぞれパキケファロサウルスの亜成体と幼体であるとも言われる。
頭部は仲間同士のディスプレイまたは実力行使、もしかしたら捕食者の撃退に用いられた。ドームは平らなものと丸く盛り上がったものに大別される。
従来、仲間との闘争で頭突きをするときは助走をつけて頭と頭を勢いよくぶつけ合うとされ、その際の衝撃は一直線に伸ばした脊椎を伝わって太い尾に分散されると考えられていた。
現在では、それだと細い頸椎に加わる負担が大きくまた丸い頭が滑ったときの危険も大きいため、頭をゆっくり押し付けあうか頭で相手の脇腹を押したとする説、頭突きは行わずディスプレイのみだったとする説が出されている。
発達した首の筋肉で衝撃を受け止めたのかもしれない。
代表的な種類
| 原始的なもの | ヤヴェルランディア、ワンナノサウルス | 頭部の特徴は未発達。 |
| ホマロケファレ類 | ゴヨケファレ、ホマロケファレ | 頭部は扁平で、丸いドームではない。 |
| パキケファロサウルス類 | ステゴケラス、ティロケファレ、パキケファロサウルス、スティギモロク、ドラコレックス■ | ドラコレックス以外は盛り上がったドームを持つ。 |
角竜類

|
| トリケラトプス |
植物食(あるいは雑食)、原始的なものとプシッタコサウルス類を除いて四足歩行。鋭いクチバシと切り刻むのに適した歯、角やフリルのある大きな頭部、頑丈でどっしりとした首や胴体・四肢、短い尾を持つ。
トリケラトプス以外のフリルには大きな穴が開いており、生前は軟組織で埋められていたとはいえ頑丈な盾というわけではなかった。角竜全てに見られる鋭いクチバシは噛み千切る力が非常に強かった。
密集して「ボーンベッド」をなす状態で見つかることがある。化石に含まれる酸素同位体の比率は雑食を示す。
またトリケラトプスも老化するとフリルに穴が開き、従来トロサウルスと呼ばれていた恐竜の姿になったのではないかとする説がある(トリケラトプスの記事参照。統合される場合名前は先に命名されたトリケラトプスが優先される)。
クチバシや歯の形態から、もしかしたら恐竜の死体を骨ごと齧ってカルシウム補給を行っていたかもしれない。
代表的な種類
| 原始的なもの | インロン、カオヤングサウルス | 二足歩行で身軽。クチバシはすでに発達。 |
| プシッタコサウルス類 | プシッタコサウルス■、ホンシャノサウルス、アルバロフォサウルス | 二足歩行。角ばった頭部にあまりとがっていない強力なクチバシを持つ。プシッタコサウルスの尾に羽毛らしき剛毛が見つかった。 |
| 原始的なケラトプス類 | リャオケラトプス、アーケオケラトプス、ウダノケラトプス、レプトケラトプス | 二足歩行ないし四足歩行。フリルを含めた頭部が大型化していった。 |
| プロトケラトプス類 | プロトケラトプス、バガケラトプス、マグニロストリス、プラティケラトプス | 四足歩行。フリルは発達しているが角はないかごく小さい。主にアジアで繁栄。 |
| セントロサウルス類 | ディアブロケラトプス、セントロサウルス、スティラコサウルス、アケロウサウルス、パキリノサウルス■ | 大半は鼻の上の一本角が発達していたが、進化するにつれ変形して骨のこぶとなった。 |
| カスモサウルス類 | カスモサウルス、ペンタケラトプス、アンキケラトプス、アリノケラトプス、トリケラトプス■ | 目の上の二本の角が発達していた。大半はフリルが縦長。 |
鳥脚類

|
| パラサウロロフス |
植物食、二足あるいは四足歩行(大型のものは場合によって前足をついた)。クチバシと発達した奥歯を持つ頭部、縦に大きい胴体と尾、長い後肢とそれ程でもない前肢を持ち、凶器や装甲の類はほとんどなかった。
膝の裏にある腱の収まる溝が恐竜の中で最も深く、これは素早く後肢を動かすのに適した形態である。
大型かつ最も派生的なランベオサウルス類は頭部に中空のトサカを持つ。これは内部が鼻につながっており、また種類によって非常に様々な形をしている。
北米の白亜紀末からエドモントサウルスなどのミイラ化石が発見されており、恐竜の中では鱗や角質、足の裏の肉球のようなふくらみなど軟組織の様子がよく分かっている。角竜同様ボーンベッドをなすことがある。
身を守るのに用いたのは、発達した後肢の脚力と、一部のものが持っていたであろう群れを成す習性だけだった。一見無防備そうだが、植物食恐竜の中では特に繁栄したグループの一つである。
トサカは発声、ディスプレイ(これらは群れをなすことと関連する)、呼気・吸気の温度調節、嗅覚増強など様々な役目が考えられる。
代表的な種類
| ピサノサウルス類 | ピサノサウルス、ファブロサウルス | 鳥脚類ではなく鳥盤類全体で最も原始的なグループだが、便宜上ここに掲載。顔つきが他の爬虫類と変わらない。 |
| ヘテロドントサウルス類 | ヘテロドントサウルス、ペゴマスタックス、フリタデンス、ティアニュロング | これも鳥脚類より原始的で、ごく小型。犬歯を持つ。ティアニュロングから羽毛らしき剛毛が見つかった。 |
| ヒプシロフォドン類 | ヒプシロフォドン、オリクトドロメウス、ドリオサウルス、レエリナサウラ、テスケロサウルス | 小型で常に二足歩行。世界中で長期間繁栄。目が大きい。 |
| イグアノドン類 | カンプトサウルス、イグアノドン、フクイサウルス、オウラノサウルス、プロバクトロサウルス | 大型・四足歩行化。奥歯が発達。前肢親指がスパイク状になっていた。白亜紀前期の各地で繁栄。 |
| ハドロサウルス類 | ハドロサウルス、エドモントサウルス、マイアサウラ、シャントゥンゴサウルス、サウロロフス | 特に大型化し、奥歯が非常に発達。トサカはないか小さな骨のでっぱり。広義にはランベオサウルス類を含む。 |
| ランベオサウルス類 | ランベオサウルス、パラサウロロフス、コリトサウルス、ニッポノサウルス、オロロティタン | ハドロサウルス類とよく似るが鼻道につながった中空のトサカが発達。 |
| 獣脚類1 2 | 竜脚形類 | 剣竜類 | 鎧竜類 | 堅頭竜類 | 角竜類 | 鳥脚類 |
恐竜以外の絶滅爬虫類
ここでは現在生き残っているものがいないグループのうち特に繁栄した、翼竜類、魚竜類、首長竜類、モササウルス類、単弓類を挙げる。
中生代にはこれ以外にワニやカメも大いに繁栄した。
翼竜類

|
| プテラノドン |
分類上恐竜に近縁。長く伸びた薬指で支えられた皮膜の翼、大きく細長い頭部、非常に華奢な骨格を持つ。歯がないと思われがちだが、小型~中型のものには歯があった。
膜の翼は一見頼りなく見えるが、膜の内部には筋肉と思われる丈夫な繊維が走っていた。これによりしわがなく破れにくい翼面を形作っていた。
ジュラ紀までは翼長1m前後の小型のものも多かったが、白亜紀に入ると大型で翼の細長いものばかりになっていった。これは鳥類が多様性を増すにつれ住み分けが進んだためと思われる。
小型のものの一部を除いて大半が魚を食べたといわれる。細かい歯やブラシ状の毛で小魚やプランクトンを濾し取ったもの、貝を割って食べたと思われるものもいた。
白亜紀末には史上最大の飛行動物・ケツァルコアトルス(翼長11m)が登場した。巨大すぎて長く羽ばたき続けるには筋肉が小さく、「飛べない」という言い方をされることもあるが、滑空でもいいから空中で生活しないと危険なほど極端な体形をしているので陸上で生活していたわけでは決してない。
こうした大型のものは自力ではなく高いところから飛び立ったと言われるが、腕の力で地表から跳ね上がったという説もある。
代表的な種類
| ランフォリンクス類 | エウディモルフォドン、ディモルフォドン、ランフォリンクス、スカフォグナトゥス、ソルデス、アヌログナトゥス、ダルウィノプテルス | アヌログナトゥス科を除いて尾が長く、小型から中型。みな歯があった。 |
| プテロダクティルス類 | プテロダクティルス、プテロダウストロ、ズンガリプテルス、タペジャラ(トゥパンダクティルス)■、ケツァルコアトルス、アンハングエラ、プテラノドン | 尾が短く、クチバシが長い。大型のものや歯がないものもいた。 |
魚竜類

|
| イクチオサウルス |
恐竜の本の挿絵に描かれた海にいたイルカみたいなやつ。と言っても後肢もひれになっており、尾びれが縦なので、首から後ろはサメのほうが似ている。
大半は数m以下の大きさ。テムノドントサウルスは8m前後、ショニサウルスは推定15mに達した。
またイルカと異なり視力が発達していたようで、大きな目を持っていた。テムノドントサウルスの眼球は脊椎動物で最大。
見た目どおり、マグロに匹敵する遊泳力を発揮して魚やアンモナイト、原始的なイカを食べたとされる。
出産時を含め水中から出ることはなかった。胎生で、腹の中に子供がいる化石も見つかっている。
海面の変化のため、または魚類の遊泳力の発達のためか白亜紀の終わりを待たずして絶滅した。
代表的な種類
| 原始的なもの | チャオフサウルス、ウタツサウルス、キンボスポンディルス | 尾びれはまだウナギのように細長く、体をくねらせて泳いだ。 |
| シャスタサウルス類 | シャスタサウルス、ショニサウルス | 尾びれが発達してくる。かなり大型化。 |
| イクチオサウルス類 | ユーリノサウルス、イクチオサウルス、テムノドントサウルス、オフタルモサウルス、プラティプテリギウス | 完全に完成した体形を持つ。 |
首長竜類

|
| プレシオサウルス |
胴体は甲羅のないウミガメと言った感じ。主に長い首と小さい頭を持ったものと、短い首と大きな頭を持ったものがいた。どちらも最大15mくらい。肩や腰の構造は陸上に上がるには耐えられないものだった。
首の長いものは小さな魚やイカ、アンモナイトの群れにゆっくり近づいたり、海の底に向かって首を降ろしコウモリダコなどを捕らえたとされる。
よく竜脚類がこれと混同されて首長竜と呼ばれるが、頸椎そのものが長く伸びて釣竿のようになっていた竜脚類と異なり首長竜は頸椎の数で長さを稼いでおり、竜脚類と違って首を曲げることができたと考えられる。しかし一時期考えられていたように、とぐろを巻いたり空中に高々ともたげることはできなかった。
頭の大きいものは素早く泳いで大きな魚やアンモナイト、他の海の爬虫類を食べたといわれる。
こちらも胎生で水中から出なかったという説が主流。「ドラえもん のび太の恐竜」では陸に上がり卵を産んだとしているが、卵の化石を見つけないことには話が始まらないのでしゃあない。
代表的な種類
| ノトサウルス類 | ノトサウルス、ケイチョウサウルス、パキプレウロサウルス | すでに長い首を持つが、四肢は単純な水かきだった。 |
| ピストサウルス類 | キマトサウルス、ピストサウルス、アウグスタサウルス | より水中生活に適応。 |
| プリオサウルス類 | マクロプラタ、ロマレオサウルス、プリオサウルス、リオプレウロドン、クロノサウルス | 特に首が短く頭が大きい。一部は非常に大型化し他の海生爬虫類も捕食したと考えられる。 |
| プレシオサウルス類 | プレシオサウルス■、クリプトクリドゥス、ムラエノサウルス | 首は胴体と同じくらい長く、頭は小さい。 |
| ポリコチルス類 | ドリコリンコプス、トリナクロメルム、ポリコチルス | 顎が細長く、首はやや短い。全長3m程度。胃の位置から頭足類の顎器が見つかったり、腹部に胎児が見つかったりしている。 |
| エラスモサウルス類 | エロマンガサウルス、フタバサウルス(フタバスズキリュウ)■、エラスモサウルス、タラソメドン、アルバートネクテス | 首が特に長く、全長の半分から2/3を占める。プリオサウルス類の分岐したものとする説もある。 |
モササウルス類

|
| モササウルス(旧) |

|
| モササウルス(新) |
ひれ状の尾を使って泳いだ。手足もひれ状になっていた。現生のトカゲやヘビと同じ嗅覚器官であるヤコブソン器官を備えていた。最大約10m。
白亜紀に入ると魚竜類が衰退し、モササウルス類が取って代わるように現れた。 これも海から出ることなく魚やアンモナイト、他の爬虫類を食べていたとされる。
アンモナイトのなかにはモササウルス類の歯型とされる穴が開いたものが見つかっている。
しかしこのような穴はモササウルス類の口の形に合わないものが多く、またカサガイという円盤型の貝が住処にするために岩に開ける穴に酷似している。噛み砕かれていないことも考えると、モササウルス類がアンモナイトを食べた証拠とはならないだろう。
上図のようにウナギのような尾びれを持っていたとするのが定説だったが、もっとも進化したモササウルス類であるプロトサウルスは尾椎が下に向かって曲がっており、サメに似た尾びれを持っていたという説があった。
さらに近年、比較的原始的であるプラテカルプスでも尾椎の屈曲が認められた。
今後はモササウルス類の一部あるいは全部の復元像が、下図のような尾びれを持った姿に変化するだろう。
代表的な種類
| アイギアロサウルス類 | アイギアロサウルス | 原始的で、おそらく指の分かれた四肢とまっすぐな尾びれを持っていた。 |
| モササウルス類 | ティロサウルス、プラテカルプス、クリダステス、グロビデンス、モササウルス、プロトサウルス | アイギアロサウルス類が海に出てかなり短期間で進化した。 |
単弓類

|
| ディメトロドン |
全て四足歩行。恐竜より前の時代に栄えていた。よく恐竜グッズにある、背びれが大きくて四足歩行の肉食のやつはこれに含まれる(ディメトロドン)。
あまりにも他の爬虫類と共通点が少ないので、今では爬虫類と呼ばれない。他の爬虫類との関係については「爬虫類:他の脊椎動物との関係」を参照。
恐竜が現れた三畳紀後期、ほぼ同時に哺乳類が現れた。その後恐竜の方が栄えていたのは、ペルム紀後期及び三畳紀中期の気候変動が単弓類や哺乳類より恐竜を含む主竜類に有利なものだったためと考えられる。
代表的な種類
| 盤竜類 | アーケオシリス、オフィアコドン、エダフォサウルス、ディメトロドン、コティロリンクス | 一見標準的な爬虫類のようだが、複数の種類の歯が生えるなどしていた。 |
| ビアルモスクス類 | ビアルモスクス、エオティタノスクス | ここから下を獣弓類という。肢がやや直立に近づく。大きな犬歯を持つ捕食者。 |
| ディノケファルス類 | アンテオサウルス、エステメノスクス、モスコプス | 頭部の発達したグループ。捕食者とカバに似た植物食者を含む。皮膚の痕跡から爬虫類にはない汗腺が見つかっている。 |
| ディキノドン類 | システケファルス、ディキノドン、カンネメイエリア、リストロサウルス、プラケリアス | 丸みを帯びた頭と胴体、太く大きな犬歯を持つ植物食者。 |
| 獣歯類 | ゴルゴノプス、イノストランケヴィア、トリナクソドン、キノグナトゥス、トリティロドン | 哺乳類にごく近い捕食者。腹部の肋骨がなくなったことから、横隔膜を持ち腹式呼吸ができたと考えられる。 |
| 翼竜類 | 魚竜類 | 首長竜類 | モササウルス類 | 単弓類 |
復元について
当然のことながら恐竜自体については化石の記録しか残っておらず、それを生痕化石と呼ばれる足跡や巣、糞などの化石、同じ地層から発掘される生物の化石、近縁な現生生物の知識で補って生前の姿や行動を推測するしかない。
しかしながら新たな化石の発掘や化石を調べる方法の進歩により、ほんの数年前の知識が一新されていくのも常である。恐竜学自体が比較的若い学問であり未踏の発掘地や未発見の化石は数多く、さらに調査研究のための手法も日進月歩の発展を見せているためである。
長い時間をかけた大きな復元の変化の例:イグアノドン

|
| イグアノドン復元三態 |
長いスパンの復元の変化を示す例として、イグアノドンを挙げたい。イグアノドンは最初に発見された恐竜の一つであり、それだけ長い間復元の変遷にさらされてきた恐竜でもある。
1822年にイギリスで歯や爪など一部が発見されたイグアノドンは、当初は図の手前のように、巨大なトカゲのような、単に大きな爬虫類として復元された。これは当時、恐竜の全身骨格は全く知られておらず、現在のトカゲやワニから推測するしかなかったためである。
1878年、ベルギーの炭鉱でイグアノドンの完全な全身骨格が30体以上発見されると、復元図も一つ奥のもののような、正確な骨格を踏まえたものに変わった。当初角とされていた小さな円錐形の骨は前肢の親指の爪とわかった。
長い後肢と短い前肢がカンガルーに似ていることもあり、背筋を垂直にして尻尾を引きずりながら二足歩行するように描かれた。
1960年代以降、骨格から活発な動物であることが明らかなデイノニクスが発見されたのをきっかけに、恐竜は活動的な生き物であるとされ、イグアノドンを始めとする大型鳥脚類も尻尾を引きずらず体を水平にして歩く姿に復元されるようになった。
足跡化石や物理演算は、図の一番奥のように、大型鳥脚類が前肢を地面について四足歩行をする場合も多かったことを示している。
比較的短期間で復元が変化した例:羽毛恐竜

|
| アンキオルニス |
素早く復元像を塗り替えた例にアンキオルニス等の羽毛恐竜がある。鳥類に近縁な恐竜に羽毛があったのではないかと70年代から言われてきたが、特に先進的な復元画家ですら小型恐竜の肌をウロコから羽毛に切り替えるのにはためらってきた。
しかし1995年、中国でシノサウロプテリクスが発見されてそのような躊躇は必要なくなった。シノサウロプテリクスの非常に保存状態の良い化石には、背筋に沿った羽毛の痕跡が認められたからだ。
羽毛ではなく皮下組織の繊維ではないかという一部の主張を押し流すように続けざまに羽毛の痕跡が発掘され、風切羽を持つヴェロキラプトルの復元画は当たり前になった。
さらに近年、そのような羽毛の痕跡を電子顕微鏡で観察した結果、メラニン色素を含む細胞「メラノソーム」の痕跡が認められた。
シノサウロプテリクスでは部分的な解析の結果羽毛が赤褐色であることが分かり、さらにアンキオルニスでは図のような赤っぽい冠羽と白黒ツートンの羽衣を持つことが分かった。
また始祖鳥も最初に発見された羽毛を観察した結果、少なくとも翼の一部は黒かったことが分かった。
恐竜の体色は知り得ないものとされてきたが、羽毛恐竜のメラニン由来の体色に関してはそれは覆されたといえる。
恐竜は恒温動物か?変温動物か?
これは研究者の多くを悩ませていた問題である。初期の頃は恐竜はすべて爬虫類として変温動物に押し込まれていた。だが、最近になって恐竜直系の子孫である鳥類が恒温動物であることからこれに疑問を呈する意見が出始めたのである。現在ではその分類により三つの意見が出されている(鳥盤類についても同等の体重のものと同じ)。
まず鳥類の直系の祖先となった小型の獣脚類。彼らはその筋肉の付き方から長時間の激しい運動に耐えられたと考えられていること、そしてその多くが体温を維持する羽毛を持っていたと考えられていることから、恒温動物であったことは確実視されている。
次は大型の竜脚類。彼らはその巨体を維持するために恒温動物同様の二心房二心室の心臓を必要としていたと考えられることと、その体重の割に体表面積はあまり大きくなく体温が逃げにくい構造になっていることからこれも恒温動物であったと考えられている。
最後にティラノサウルスに代表される大型獣脚類であるが、これは現在でも意見が別れている。ある学者は呼吸器がその巨体を長時間激しく運動させるには不十分であるとして変温動物説を主張し、別の学者は筋肉の付き方から恒温動物であると主張している。
最近の研究では大型獣脚類の骨格から鳥類にも見られる気嚢という呼吸器官の痕跡が見つかり、実は呼吸効率は思ったより高かったこと、そして全身に分布した気嚢により大型獣脚類の体内は実はかなりの部分が空洞で従来の見積もりより体重が軽く、長時間の運動に耐えられた可能性があることから恒温動物説側に意見が傾きつつある。
伝熱工学的に見ると、大型の動物ほど体温維持に費やす自分自身の代謝エネルギーは少なくて済む。実際にワニの体温を測定したところ、変温とされるワニでも大型なら体温の変化が少ないことが判明した。大型の恐竜が体温を一定に保っていたとしても、自分自身ではほとんど熱を生み出す必要がなく食糧確保に悩まなくてよかっただろうと考えられる。このような生理を慣性恒温という。
しかし、完全に慣性恒温に頼っているワニでは同程度の体重の恒温動物に対して筋力や持続力で大幅に劣るという研究結果もある。このことが問題になるとすれば、恐竜の場合はわずかでも自分自身の代謝で体温を保っていただろう。
骨組織の様子から成長の速度を読み取ると、恐竜の成長は変温動物と恒温動物の中間で、一部のサメやマグロ、オサガメのような、基本的には変温動物だが体の熱を逃がさないことで体温を保つ仕組みを持つ動物に近いという。
絶滅について
1億6千万年もの間陸上生態系に君臨してきたにもかかわらず、恐竜は鳥類以外全て絶滅してしまった。これは一体なぜなのか、その原因は人々の興味をひき、病気の蔓延・食中毒・超新星爆発による放射線・部長の打ち返し等様々な推測を呼んだ。
恐竜が絶滅したのは現在から6550万年前の白亜紀後期である。このときの地層を調査すると、通常地殻には現れないイリジウムの濃度が高いことがわかった。またメキシコ・ユカタン半島沖に、恐竜絶滅当時に小惑星が衝突してできたクレーターが発見された。
しかしそれ以外にも火山活動の活発化、生態系の変化など様々なことが白亜紀末に起こったことが分かっており、そちらに絶滅の原因を求める反論も根強かった。
そこで植物や菌の花粉・胞子、プランクトン殻、各種動物の化石、イリジウムに限らず衝撃変成石英などの堆積物等の様子を詳細に調査することで、それらの反論に対する再反論の材料が集められ、小惑星衝突により白亜紀末の大絶滅が起きたことはほとんど確実といえるようになった。
この間、衝突原因説は定説としての地位を確立していったが、メディアに取り上げられるのは「定説を覆す研究」としてよりセンセーショナルに扱える反論のほうであった。
そしてこのような状況を受けて、2010年3月5日、各分野の41人の共著者によって「小惑星衝突が原因である」と宣言する論文が「サイエンス」誌に掲載された。 この経緯は下記の書籍に分かりやすくまとめられている。
ただ注意してほしいのは、絶滅したからと言って恐竜に生き物として根本的な欠陥はなかったということ(化石がたくさん残っていることと、それまで哺乳類と共存して長いこと繁栄してきたのが証拠)。そして、恐竜の全てが絶滅したのではないということである。
公園や路上、ごく身近なところでも恐竜達は今も繁栄している。羽毛を持った小さな恐竜達を見かけたら、是非彼らの辿ってきた歴史、彼らが選んだ進化の道に思いをはせてみてほしい。
命名について
恐竜をはじめ古生物の呼び名は、他の生き物にも付けられる世界共通の名前「学名」である。
一般的には属名(学名の前半部分。ごく小さなグループの名前)だけで呼ぶが、専門的には種小名(後半部分。属名と合わせて単一の種類を表す)も必要である。
例えばティラノサウルス属はレックス種1種のみだが、ステゴサウルス属にはアルマトゥス種、ステノプス種、ロンギスピヌス種、ウンギュラトゥス種などが含まれる。今の生き物で言うと、パンテラ属にレオ種(ライオン)、オンカ種(ジャガー)、パルドゥス種(ヒョウ)、ティグリス種(トラ)が含まれるのと同じことである。
学名はラテン語かラテン語化したギリシャ語で命名されるのだが、人名や地名、団体名、現地語など他の言語をラテン語の造語として用いることもできる。
化石の特徴や推定される生態、発見された土地や発見に関わった人物の名前等に基づいて名付けられることが多い。
まず、よく用いられる単語の一部を例に挙げると、意味は以下のとおり。
- サウルス(saurus):トカゲ、爬虫類 →サウロ(sauro-):トカゲの
- ラプトル(raptor):強盗、猛禽
- ティタン(titan):巨人 →ティタノ(titano-):巨大な
- ミムス(mimus):もどき
- オルニス(ornis):鳥 →オルニト(ornitho-):鳥の
- ケラス(ceras):角 →ケラト(cerato-):角の
- ロフス(lophus):トサカ →ロフォ(lopho-):トサカの
- (オ)プス(-ops):顔面
- プテラ(ptera)、プテリクス(pteryx):羽、翼 →プテロ(ptero-):羽の、翼の
- (オ)ニクス(-onyx):爪
- (オ)ドン(-odon):歯
- デイノ(deino-):恐ろしい ディノ(dino-):恐ろしく大きな
例えば以下のような意味で名付けられている。
- 化石の特徴によるもの
ケラトサウルス(Ceratosaurus 角の生えたトカゲ)、コンプソグナトゥス(Compsognathus 華奢な顎)、ブラキオサウルス(Brachiosaurus (脚より長い)腕のトカゲ)、トリケラトプス(Triceratops 三本角の顔)、イグアノドン(Iguanodon イグアナのような歯)など - 推定される生態によるもの
トルヴォサウルス(Torvosaurus 残虐なトカゲ)、ティラノサウルス(Tyrannnosaurus 暴君トカゲ)、ヴェロキラプトル(Velociraptor 敏捷な強盗)、オロドロメウス(Orodromeus 山で走るもの)、マイアサウラ(Maiasaura 良い母のトカゲ)など - 分類や他の恐竜との比較によるもの
アロサウルス(Allosaurus (脊椎の構造がそれまで見つかっていた恐竜と)異なるトカゲ)、ファルカリウス(Falcarius 鎌職人(=テリジノサウルス(「大鎌トカゲ」)の祖先))、アンテトニトルス(Antetonitrus 雷(=雷竜、つまり竜脚類)の前)、プロトケラトプス(Protoceratops 原始的な角竜)、パラサウロロフス(Parasaurolophus 副サウロロフス(サウロロフスは「トサカのあるトカゲ」))など - 伝説に基づくもの
ガルディミムス(Garudimimus 神鳥ガルーダに似たもの)、ジョバリア(Jobaria 怪物ジョバール)、サウロポセイドン(Sauroposeidon 地震を起こす海神のトカゲ)、スティギモロク(Stygimoloch 三途の川の鬼神)、ガーゴイロサウルス(Gargoyleosaurus 石像の化身トカゲ)など - 土地や人物などによるもの(「献名」と言い、基本的に命名者自身の名前は使わない)
フクイラプトル(Fukuiraptor 福井の強盗)、シノヴェナトル(Sinovenator 中国の狩人)、アルゼンチノサウルス(Argentinosaurus アルゼンチンのトカゲ)、ガストニア(Gastonia ロバート・ガストン氏のもの)、アトラスコプコサウルス(Atlascopcosaurus アトラスコプコ社が発掘に協力したトカゲ)など
最も長い名前の恐竜はミクロパキケファロサウルス・ホングチュヤネンシス(Micropachycephalosaurus hongtuyanensis 紅土岩で発見された小さくて頭の分厚いトカゲ)で、最も短い名前の恐竜はイー・チー(Yi qi おかしな翼)である。
学名はその生き物を最初に発表した論文で名付けることができ、これを記載という。いくつかの命名上のルールがあるが、「研究が進んで分類を変更する際は、先に名付けられた方の名前が優先される」という大原則がある。
例えば「大腿骨の化石A」と「頭骨の化石B」が見つかって別々に研究され、各々独自の特徴が認められたためAが「ニコドウサウルス」、Bが「ニコヒャクケラトプス」と命名されたとする。
もし後に「Aそっくりの大腿骨とBそっくりの頭骨を含むほぼ全身の化石C」が発見され、AとBが同じ種類の恐竜のものであると判明した場合、「ニコドウサウルス」と「ニコヒャクケラトプス」のうち先に論文が発表された方の名前を使わなければならない。「ニコドウサウルス」の論文が2007年1月15日、「ニコヒャクケラトプス」の論文が2008年5月12日に発表されたとしたら、ニコヒャクケラトプスと呼んでいたものもニコドウサウルスと呼ばなくてはならない。
また、さらに別の化石Dに「ニコナマラプトル」と名付けたが、ニコナマラプトルという名前がすでに別の生き物に付けられていたという場合、また別の名前を付け直さなくてはならない。現生種と名前がかぶることもよくある。
こちらの動画では「エロマンガサウルス」という首長竜の命名についてUTAU音声で解説している。
元ネタのある恐竜・古生物の名前
古生物の名前の中には文学やサブカルチャーから取ったものまである。例えばこんな具合。
- ゴジラサウルス・クエイイ
三畳紀後期北米の肉食恐竜。全長5.5mと当時としては大型なことから、怪獣映画ファンであるケネス・カーペンターによって命名された。綴りは英語版に基づくGodzillaではなく発音に忠実なGojiraである。残念ながら現在ではコエロフィシス属に含まれ無効名とされるようだ。 - ボロゴヴィア・グラキリクルス
白亜紀後期モンゴルの小型肉食恐竜。「不思議の国のアリス」の劇中詩「ジャバウォッキー」に登場する鳥ボロゴーヴにちなんでハルシュカ・オズムルスカが命名。足しか見つかっていないが名前どおり鳥に近い種類だった。 - ドラコレックス・ホグワーツィア
白亜紀後期北米の堅頭竜類。名前は「ホグワーツ魔法学校の竜王」を意味する。角や棘でびっしりと覆われた頭骨があまりにもドラゴン然としていることから、「ハリー・ポッター」シリーズにちなんでロバート・バッカーが命名。パキケファロサウルスの幼体と考えられることが多い。 - インロング・ドウンシ
ジュラ紀後期の原始的な角竜。属名は中国語で「隠竜」と書き、発掘現場近くで香港映画「グリーンデスティニー」を撮影していたことから、その原題である「臥虎蔵龍」の「蔵龍」と同じ意味を持たせて徐星が命名。翼竜のクリプトドラコンは同じ由来で名付けられた。 - ティアンチサウルス・ネデゴアペフェリマ(ジュラッソサウルス・ネデゴアペフェルキモルム)
ジュラ紀中期中国の鎧竜類。改名前の名前を無理矢理意訳すると「ジュラシックパークに出演したみんなのトカゲ」。種小名(名前の後半)はジュラシックパークの出演者の頭文字をつなげたもので、スティーブン・スピルバーグが中国科学院古脊椎動物・古人類学研究所に研究資金を寄贈した見返りに董枝明に命名させた。 - クリトンサウルス・ボーリニ
白亜紀前期中国の鎧竜類。クリトンはジュラシックパークの原作者マイケル・クライトンのこと。こちらも董枝明の命名。種小名ボーリニは北京原人を研究したスウェーデン人研究者ボーリンへの献名。 - シネミス・ガメラ
白亜紀前期中国のカメ。甲長20cmほどの、今のイシガメなどとさして変わらないカメだったが、六角形の甲羅には後ろ側左右から飛行機の翼のような突起が伸びていた。このため「ステルスボンバータートル」というあだ名がつき、それを受けてドナルド・ブリングマンとペンが命名。 - ニンジェミス・オウエニ
更新世オーストラリアのリクガメ。後頭部から伸びた二つの角を忍者装束の頭巾に見立てて、「ニンジャ・タートルズ」にちなんでギャフニーが命名。 - レヴィアタン・メルヴィレイ
中新世のマッコウクジラの仲間。レヴィアタンはリヴァイアサンのラテン読み。頑丈な顎と巨大な牙があり今のマッコウクジラと比べかなり捕食傾向が強いと考えられることから、ハーマン・メルヴィルの「白鯨」にちなんでオリビエ・ランベールが命名。
恐竜を元ネタとするもの
特撮モノ
ゴジラシリーズ(ゴジラ、アンギラス、バラゴンなど)、ウルトラマンシリーズ(レッドキング、ゴモラなど)が特に有名。キングコングにおけるティラノサウルスや日本版キングコングにおけるゴロザウルスなど恐竜でありつつ怪獣として振る舞うものもあり、かつては映画での扱いに明確な境界線がなかったようだ。
尻尾を引きずった旧復元スタイルにのっとった怪獣の姿は日本人の恐竜観に大きな影響を与えた。現在でも恐竜が「ゴジラ立ち」をしていたと思っている人や、恐竜と怪獣の区別がよく分からない人もいるようだ。
怪獣を主人公とした映画としてゴジラと並び立つガメラシリーズには明確に恐竜など古生物をモチーフとしたものは登場しないが、平成ガメラシリーズの世界ではカメ類は恐竜とともに絶滅したものとされている。
また、ヒーロー側のモチーフとして恐竜が扱われる事も多く、スーパー戦隊シリーズではこれまで四作(恐竜戦隊ジュウレンジャー、爆竜戦隊アバレンジャー、獣電戦隊キョウリュウジャー、騎士竜戦隊リュウソウジャー)登場している。
ジュラシックパークシリーズ
「恐竜ルネッサンス」以降の活発な恐竜像を当時の最新映像技術で克明に描いた、恐竜映画の金字塔として名高い。恐竜と人類が出会う方法に科学的な説得力を持たせたことも印象を深めている。
恐竜ファンとして見ると功罪相半ばすといったところで、知識がないと画面の中では本当に科学的な描写とエンターテインメントとしての脚色が同じように見えるため混乱を招いた面がある。最も誤解されたのは、映画では大型種ユタラプトルに近い姿で描かれたヴェロキラプトルであろう。(もっとも作中に登場する恐竜はDNAを基にしたクローンなので現実の学術的に矛盾があってもさほど問題がないように作られているが)
映画独自の設定や描写が後の恐竜グッズやイラスト等でも当然のように引用されるくらい影響力があり、よく見るとティラノサウルスのフィギュアがみんなジュラシックパーク風だったりもする。
ファンタジー作品におけるドラゴン
大元の伝説や中世に描かれた挿絵の中ではトカゲ・ワニ・ヘビなどを基にした奇怪な姿であったが、キリスト教における悪魔の象徴であるといった宗教的な意味合いが薄れ、さらに古生物に関する知識が広まるにつれ、恐竜、特に獣脚類の要素を取り入れるようになった。
最も顕著なのが「モンスターハンター」シリーズで、二足歩行を行う種類の側面シルエットなどはかなり恐竜を踏襲している。恐竜そのものだと思われることも多いほどだが、やはりゲームのキャラクターとして魅力的に描く上で実際の恐竜にとらわれない描写や設定をしている部分が多い。例えばモンハンのモンスターは大きさの割に身軽そうな勇ましいポーズを保っており、頭骨もトカゲか肉食哺乳類のように幅広い。ティガレックスやイビルジョーとティラノサウルス(特に骨格)を見比べてみよう。
ゾイドシリーズ
機械化しているという時点で充分キャラクターとしての魅力を付け加えられているためか、シルエットは原型の恐竜にかなり忠実なものも多い。ただしティラノサウルス型に限っては大半がなぜか小顔でシャープなモデル体型に。ちなみに「ウネンラギア」など元の恐竜の名前そのままのものもある。
30年近く続く歴史の中で恐竜像も変化しているため、歴代の機体にその痕跡が残っているのが興味深い。水平姿勢を取れるようになったゴジュラスシリーズ、原型の復元像が大きく変わってしまったデスレイザーやバイオメガラプトル、原型が命名にひと悶着あった上結局抹消されてしまったウルトラザウルスなど。
ポケモン
生き物だけでなく何でもかんでもモチーフにしているだけに恐竜型・古生物型ポケモンもいる。ゲーム中に化石や博物館が登場するのも魅力か。ただ特定の恐竜を直接モデルにしているものは少ない。傾向は以下の三つに分かれる(番号順、最終進化形のみ)。
恐竜型怪獣の流れを汲むもの:ニドキング、バンギラス、ボスゴドラ、ガブリアス、オノノクス、ヌメルゴンなど
似ている古生物を挙げられるもの:ラプラス(プレシオサウルス)、メガニウム(カマラサウルス)、ジュカイン(デイノニクス)、トロピウス(ディプロドクス)など
明らかに古生物や「生きた化石」をモチーフとしているもの:オムスター(アンモナイト)、カブトプス(カブトガニ+ウミサソリ+三葉虫)、プテラ(ランフォリンクス)、ユレイドル(ウミユリ)、アーマルド(アノマロカリス+ウミサソリ)、ジーランス(シーラカンス)、ラムパルド(パキケファロサウルス)、トリデプス(角竜)、メガヤンマ(メガネウラ)、マンムー(マンモス)、アバゴーラ(アーケロン)、アーケオス(始祖鳥)、ガチゴラス(ティラノサウルス)、アマルルガ(アマルガサウルス)
実際に見学するにあたって
この記事で紹介しているような恐竜の姿や生態に関する知識も、全ては化石から少しずつ得られたものである。そのため恐竜ファンとしても博物館やイベントで骨格や化石を見て、恐竜に関する説の根拠を確認するのが望ましい。またそんな向学的な目的でなくても、恐竜の大きさやスタイリングを実際に見て感じ、楽しみたいと思うのは自然だろう。
そこでここには博物館や恐竜博・恐竜展で骨格や化石を見学する際の諸注意を挙げておきたい。
レプリカと実物化石、復元骨格
「レプリカ」「複製」といった表記を見て「なんだ偽物か」という冷めたリアクションを取る見学者は後を絶たない。これらの言葉から美術品における贋作、複製品を思い起こし、作り物、張りぼて、といった無価値な物と混同してしまうのだろう。
しかし複製化石を作成する際は実物から精密に型を取り、化石の形状が完全に再現されるよう技術を凝らしている。そのため、内部の組織などを除いた形態などは実物と全く同じといってよく、形態学的な観察には何の問題もない。
また型を取った元の化石が発掘された場所や状況、所蔵している博物館が記録されているものは、実際の研究にも充分有用な学術的価値のある立派な標本である。これにより実物を破損・紛失することや、一つの研究機関で独占してしまうことを防げる。
複製は実物化石と異なりプラスチックやFRPで作ることができるので、実物よりずっと軽く丈夫で、多少加工しても問題ない。そのため支柱を内部に通して復元骨格を作るのもたやすく、非常に観察しやすいものになる。博物館で一般来館者が様々な恐竜の復元骨格を見られるのも複製技術のおかげであるといえる。
ほとんどの恐竜はごく一部の化石しか見つかっていないので、骨格を復元するにも他の大部分見つかっている種類を参考にする必要がある。どの部分がすでに発見されていてどの部分が推測なのか把握しておくとよい。一見しただけでどこが推測か分かる場合もある(表面が滑らか、色が違う、そこだけ他の種類に似すぎている、逆に近縁種と違いすぎる、なんかわざとらしい、作りが雑、等)。全身が判明していて研究の進んだ種類で一般的な形態を見て、変わった特徴を持つものについてはそこを重点的に見るのがいいだろう。
骨格全体が複製ではなく模型として作られた骨格復元模型もある。地層中で押しつぶされて板状になった化石など立体的な複製が作れない場合などもあるので、これも研究成果をまとまった形で見て元の生き物の姿を知るのに有用である。
もちろん、実物化石には実際に化石化してできた成分、複製しきれない微細構造、また本当に悠久の時間を越えてきた事実というものがある。そういったものを観察し、また肌で感じるには実物化石が必要だろう。
実物化石は復元骨格にせずに部分状態でケースに入ったり台に置かれている場合が多いので、そういったものにも気を付けて見学しよう。
またごく一部、ほとんど実物化石で組まれた復元骨格もある。扱いに融通が利かないデメリットはあるが、相応の迫力が感じられるだろう。
種類を見分けよう
大型肉食恐竜を見て種類に関わらずティラノサウルスと呼ぶ見学者、というのもまたありがちな光景である。もちろん上記のように大型肉食恐竜は決してティラノサウルスだけではなく、多様な分類・分布・時代に及ぶ種類がある。
せっかくなので恐竜の多様性をきちんと理解して見学するといいだろう。それには大まかな違いを把握しておく必要がある。
例えばティラノサウルス類は後頭部の幅が広く前肢が二本指で小さいが、アロサウルス類は後頭部の幅があまり広くはなくて前肢がより大きい、など。
ティラノサウルスとタルボサウルスのように見た目ではほとんど区別できないこともあるので、何の仲間かだけパッと見で区別してから解説を読んで、それから特徴を確認する程度でも問題はない。またあらかじめ何が展示されているのか公式サイト等で確認するのも、目当ての恐竜があるか、何を楽しみにして行くべきかが分かって良い。
骨学的に見よう
「逆関節」という言葉がある。二足歩行をするものの脚の中央にある関節が人間の膝と逆に曲がるということで、架空兵器ではダチョウなどを参考にしたデザインである。しかし、ダチョウ自身の膝は実は「逆関節」ではない。
ダチョウの本当の膝は胴体の脇にぴったりと寄せられて羽毛に隠れている。逆に曲がった膝のような関節は人間でいえばかかと、脛に見えているのは長い足の甲に当たる。恐竜もこのような爪先立ちに近い歩き方、または完全に指だけを接地する歩き方をしていた(かかとが地面に着く人間のほうが少数派である)。
このようにどこが何の骨なのか、さらに骨のどの部分にどんな機能があったかを理解していると、どのような動作をしたか、どのくらい速く、また強く、広く動かせたか、といったことが理解しやすい。また関節の様子を見ることで、無理のない繋がり方をした正確な復元であるかどうかも確かめることができる。無理に骨の名前などを頭に叩き込まなくても個々の恐竜の特徴を知る過程で覚えられるだろう。
復元に関する書籍はとても参考になる。
イベントにおいて
どの恐竜博・恐竜展でも「目玉」としてプッシュされている復元骨格がある。大型竜脚類をその位置に置くことが多いが、大型竜脚類はほとんど一部しか見つからないものであり上記のように推定の部分がほとんどにならざるを得ない。竜脚類に限らず見世物的要素の強い復元骨格も出てくることがあるので、一緒に展示されている実物化石で大きさの復元が妥当かなど確認するといい。
また宣伝の上ではあまり取り上げられない、ガラスケース内にある実物化石のほうによっぽど貴重で保存状態も素晴らしいものが潜んでいることはよくある。他の見学者が骨格に見とれている隙にそちらもしっかりと見ておこう。
うっかり間違った解説などがなされている場合もあり、注意が必要である。具体的な例を一つ挙げると、「世界の巨大恐竜博2006」において獣脚類シノヴェナトルとして展示されていた化石の頭部には明らかに鳥脚類の特徴であるクチバシや眉毛状の骨があった。
恐竜に関わる仕事をしている人のブログなどでイベントレポートを読むと、見所を教えてもらえる上間違いも回避しやすいだろう。
見学動画
恐竜全体に関する教養動画、博物館やイベントを見学したレポート動画もあり、展示品の種類や館内の雰囲気、見方のコツなど見学する際の参考になるだろう。
律っちゃんの恐竜講座/がんびっとP
恐竜の骨格標本画像を集めてみた(種類ごと、世界各地)
日本を代表する恐竜博物館、福井県立恐竜博物館の見学動画
雪歩とめぐる大恐竜展/パキケファロサウルスP
関連項目

|
| ティラノサウルス旧復元図 |
- 動物の一覧
- バージェス動物群(カンブリアモンスター)
- ワニ(恐竜に近縁)
- 鳥
- 爬虫類
- 動物・植物・風景のお絵カキコ
- 地球 / 地球科学
- 絶滅
- 恐竜惑星
- ダイナソー
- 時をかけるおっさんシリーズ
- 化石








